
Effects of grazing and climate change on sandy grassland ecosystems in Inner Mongolia
2011-12-09 07:12:38HaLinZhaoToshiyaOkuroRuiLianZhouYuLinLiXiaoAnZuo
HaLin Zhao , Toshiya Okuro , RuiLian Zhou , YuLin Li , XiaoAn Zuo
1. Cold and Arid Regions Environmental and Engineering Research Institute, Lanzhou, Gansu 730000, China 2. Graduate School of Agricultural and Sciences, University of Tokyo, Tokyo 1138657, Japan 3. Faculty of Life Sciences, Ludong University, Yantai, Shandong 264025, China
Effects of grazing and climate change on sandy grassland ecosystems in Inner Mongolia
HaLin Zhao1*, Toshiya Okuro2, RuiLian Zhou3, YuLin Li1, XiaoAn Zuo1
1. Cold and Arid Regions Environmental and Engineering Research Institute, Lanzhou, Gansu 730000, China 2. Graduate School of Agricultural and Sciences, University of Tokyo, Tokyo 1138657, Japan 3. Faculty of Life Sciences, Ludong University, Yantai, Shandong 264025, China
To understand the effects of grazing activities and climate change on sandy grassland ecosystems in northern China, a livestock field grazing and enclosure experiment was conducted from 1992 to 2006 in Horqin Sand Land, Inner Mongolia. The results showed that sustained heavy grazing resulted in serious degradation of the vegetation; moderate grazing can maintain vegetation stabilization; and light grazing can promote rapid restoration of degraded vegetation. The livestock productivity was the highest in the moderate grazing grassland, and sustained heavy grazing resulted in rapid decrease of the livestock productivity. Heavy grazing can cause a retrogressive succession of grassland vegetation, whereas moderate and light grazing may promote progressive succession of plant species. The effects of changing climate on succession processes were not significant in the short term; a warm-humid climate is favorable to restoration of degraded vegetation, whereas a sustained warm-drought climate may result in degradation of grassland vegetation. Heavy livestock grazing should be stopped for the sustainable use of grassland; the proper grazing intensity for sandy grassland is two to three sheep or sheep equivalents per hectare in Inner Mongolia.
grazing intensity; climate change; sandy grassland; ecosystem dynamics; Inner Mongolia
1. Introduction
Grazing activities and climate change are considered to be among the primary factors that influence grassland ecosystems worldwide (Schultzet al., 1995; Nautiyalet al.,2004). In recent decades, with the rapid increase of world population and marked changes in global climate (Geet al.,2007), the effects of grazing activity and climate change on grassland ecosystems have resulted in marked degradation of grassland ecosystem in most areas of the world (Giourgaet al., 1998; Liet al., 2006). To control grassland degradation and increase grassland productivity, certain measures,including decreasing grazing intensity, fencing degraded grassland, planting perennial grasses, and applying fertilizer have been implemented in many countries in recent years(Zhaoet al., 2003; Geet al., 2007). Studies concerning the effects of grazing and climate change on grassland vegetation or grassland ecosystems are gaining increasing attention(Carmel and Kadmon, 1999; Zhaoet al., 2004; Geet al., 2007).
There is a broad literature concerning the effects of grazing management and climate change on grassland ecosystems (Jutila, 1999), include the impacts of sustained grazing on plant diversity and composition (Anderson and Hoffman, 2007); the effects of grazing intensity on above-ground biomass and vegetation structures (Donget al.,2002); the effects of grazing pressure on succession process and productivity (Giourgaet al., 1998); desertification processes of sandy rangeland due to overgrazing (Zhaoet al.,2005); the effects of enclosure (fencing) on vegetation recovery in desertified grassland (Zhenget al., 2005); the responses of grassland plants to grazing and protection(Noy-Meiret al., 1989); the effects of grazing and climatic changes on grassland vegetation (Nautiyalet al., 2004;Augustine and McNaughton, 2006); and global analysis of the relationships among a vegetation index, precipitation,and land-surface temperature (Schultz and Halpert, 1995).These studies are of great importance to understand the processes and mechanisms of the influences of grazing and climate change on grassland ecosystems, and to develop corresponding measures for grassland management (Donget al., 2002; Cooperet al., 2005). However, information is still lacking, especially on the synergistic effects of grazing, enclosure and changing climate on grassland vegetation (Nautiyalet al., 2004; Augustine and McNaughton, 2006).
The Inner Mongolia steppe is one of the largest and most important steppes in the world, also one of the most severely degraded grasslands in northern China (Zhaoet al., 2004).In recent years, continual drought has aggravated the grassland degradation processes (Liet al., 2006; Liu and Li,2006), but thus far there have been few studies concerning the synergistic effects of grazing and climate change on grassland ecosystem in this region (Liet al., 2006; Liu and Li, 2006). Therefore, the main objectives of this study were(1) to compare the influence of different grazing intensities on plant species diversity, canopy coverage, height, aboveand below-ground biomass, and livestock productivity in the grassland; (2) to analyze restoration and succession processes of grassland vegetation which were influenced by their original states and by changing climate after removing graze;and (3) to evaluate the relationships during the ecosystem changes, the grazing intensity, and climatic changes.
2. Materials and methods
2.1. Study area
The study area is located in Naiman County (42°55′N,120°42′E, 345 m elevation) in the eastern part of Inner Mongolia. Naiman County is located in the hinterland of Horqin Sand Land, it is in the continental semi-arid monsoon climate area of the temperate zone. The mean annual precipitation is 353.2 mm, the mean annual potential evaporation is 1,935 mm, and the mean annual temperature is 6.9°C. The annual frost-free period is about 130 to 150 days.The average annual wind speed is 3.4 m/s and the mean wind speed at a height of 2 m above the ground in spring is 4.3 m/s. The geomorphology in this region is characterized by dunes alternating with gently undulating lowland areas.The soils are identified as degraded sandy Chestnut soils according to the Chinese Soil Classification System, which are mostly equivalent to the Orthi-Sandic Entisols of sand origin in terms of the FAO-UNESCO system (Zhaoet al.,2003). The thickness of the soil layer in the grassland is about 15 to 30 cm, and the soil is mainly consisted of coarse sand and silt. The main plant species arePennisetum centrasiaticum,Phragmites communis,Setaria viridis,Chloris virgata,Cleistogenes squarrosa, andAristida adscensionis(Zhaoet al.,2005).
2.2. Experimental design
The experiments were conducted during the years 1992 to 2006. A flat and open natural grassland was selected in the summer of 1991. The grassland is part of an area that has been monitored for a long period by the Naiman Desertification Research Station, Chinese Ecosystem Research Network (NDRS-CERN). The mean pre-grazing intensity for the area was 4.0 to 4.5 sheep/ha (Zhaoet al., 2003), and the grassland was experiencing slight degradation due to overgrazing by sheep, according to the classification criteria of grassland degradation degree of Zhu and Chen (1994). The site was fenced into three 1.5-ha (200m×75m) plots.
The experiment was divided into two stages. The first stage was the grazing experiment in 1992–1996, aimed at studying the effects of domestic animal grazing activity on the grassland ecosystem. The experiment design contained three grazing treatments: heavy grazing (HG, 6 sheep/ha),moderate grazing (MG, 4 sheep/ha), and light grazing (LG, 2 sheep/ha). The sheep were kept on the plots in the daytime and in the shed at night, and the same sheep were kept on the same plot each day during the experiment period from June 1 to September 30 each year. The second stage was a restoration experiment in 1997–2006. The livestock were removed from all grazing plots after finishing the grazing experiment to study the effects of climate change on the restoration and succession processes of the vegetation.
2.3. Data collection and analysis
During the experiment period, three fixed parallel sample belts were set in each plot. Each sample belt was 200 m long and 1 m wide, and the space between each sample belt was about 15 m. Twenty fixed quadrants (1m×1m) with a 10-m interval were established in each belt in August each year for determination of plant species abundance, species composition, vegetation canopy coverage (summed coverage of all species), and canopy height. At the same time, six quadrants (1m×1m) were randomly selected per plot to investigate above- and below-ground biomass. The above-ground biomass (standing biomass) was measured with the clipping method (all green parts 1 cm above the ground inside the quadrants were cut). The biomass was put into paper bags in the field, oven-dried at 85 °C for 24 hrs, and weighed in the laboratory. The below-ground biomass was measured with the soil core and washing method. Soil cores were taken in the centre of each quadrant at the depths of 0–10, 10–20, and 20–30 cm with an auger. The soil cores were washed through a 0.25-mm mesh screen for root biomass measurements. The root samples were later oven-dried at 85 °C for 24 hrs and weighed. Vegetation cover was visually estimated. Canopy height was mean height of the plants inside the quadrants. The climatic data were collected from the NDRS-CERN weather station 1,200 m away from the experiment grassland.
Species richness was calculated as the average number of species per block. Plant diversity (H) was calculated as the Shannon-Wiener index (Zhang 1998):
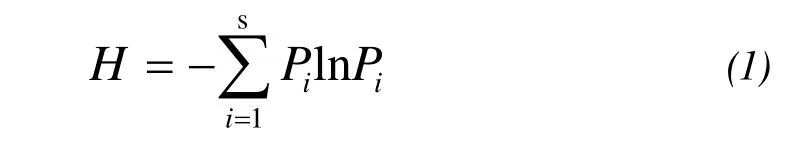
whereSis the number of species andPiis the proportional important value of theith species for each plot.
All data were analyzed using the SPSS program for Windows, version 11.5. Multiple comparison and one-way analysis of variance (ANOVA) procedures were used to compare the differences among the treatments. Least significant difference (LSD) tests were performed to determine the significance of treatment means atP<0.05 (Sokal and Rohlf, 1995).Pearson correlation coefficients were used to evaluate relationships between the corresponding variables (Zhang, 1998).
3. Results
3.1. Changes in precipitation and air temperature
Annual mean precipitate and annual mean air temperature were 325.4 mm and 7.6 °C in 1992–2006, as compared to mean values of 353.2 mm and 6.9 °C in the past 50 years.Figure 1 shows that, the precipitation was higher in 1992–1999 and lower in 2000–2006, meanwhile the temperature fluctuated in the same periods. The mean precipitation and mean temperature were 383.9 mm and 7.6 °C in the former period, and 258.6 mm and 7.6 °C in the latter, respectively. Precipitation was just 213.1 mm in 2000 and 155.6 mm in 2001, the driest stage in study period. Thus, the climate conditions in the experiment period can be divided into a warm-humid period (1992–1999) and a warm-drought period (2000–2006).
3.2. Changes of plant species richness and diversity
During the grazing experiment, species richness and diversity increased significantly (P<0.05) in the MG and LG sites, and decreased significantly (P<0.05) in the HG sites(Figure 2a). In the first two years of the restoration experiment, although the species richness and the diversity increased in all the treatments (Figure 2b), there were significant differences in the increased range among them (P<0.05). The increase ranges were the greatest in the HG sites and the lowest in the LG sites. Later, the richness and the diversity exhibited fluctuating slight decreases in all the treatments.
3.3. Changes in canopy coverage and height
During the grazing experiment, the canopy coverage and height increased significantly (P<0.05) in the LG sites, decreased significantly (P<0.05) in the HG sites, and slightly decreased (P>0.05) in the MG sites (Figure 3). The canopy coverage and height in all the treatments increased significantly in the first several years of the restoration experiment.The order of the increase range was HG sites > MG sites >LG sites. The canopy coverage and the height in all the sites presented significant fluctuating decreases (P<0.05) since 1999, and the decrease range was greater in the canopy coverage than in the height.
3.4. Changes in above- and below-ground biomass
During the grazing experiment, the above-ground biomass (standing biomass) decreased significantly (P<0.05) in both the HG and MG sites, and increased significantly (P<0.05) in the LG sites (Figure 4a). The above-ground biomass at the end of the grazing experiment was 3.3, 67.1, and 218.2 g/m2, respectively in the HG, MG and LG sites. In the early period of the restoration experiment, the above-ground biomass increased significantly in the HG and MG sites (P<0.05), and presented fluctuating change (P>0.05) in the LG sites. The below-ground biomass in the three treatments decreased significantly in the grazing experiment and increased significantly in the restoration experiment (P<0.05)(Figure 4b). The roots/stems ratios in the three treatments were significantly higher in the grazing experiment than in the restoration experiment (Figure 4c).
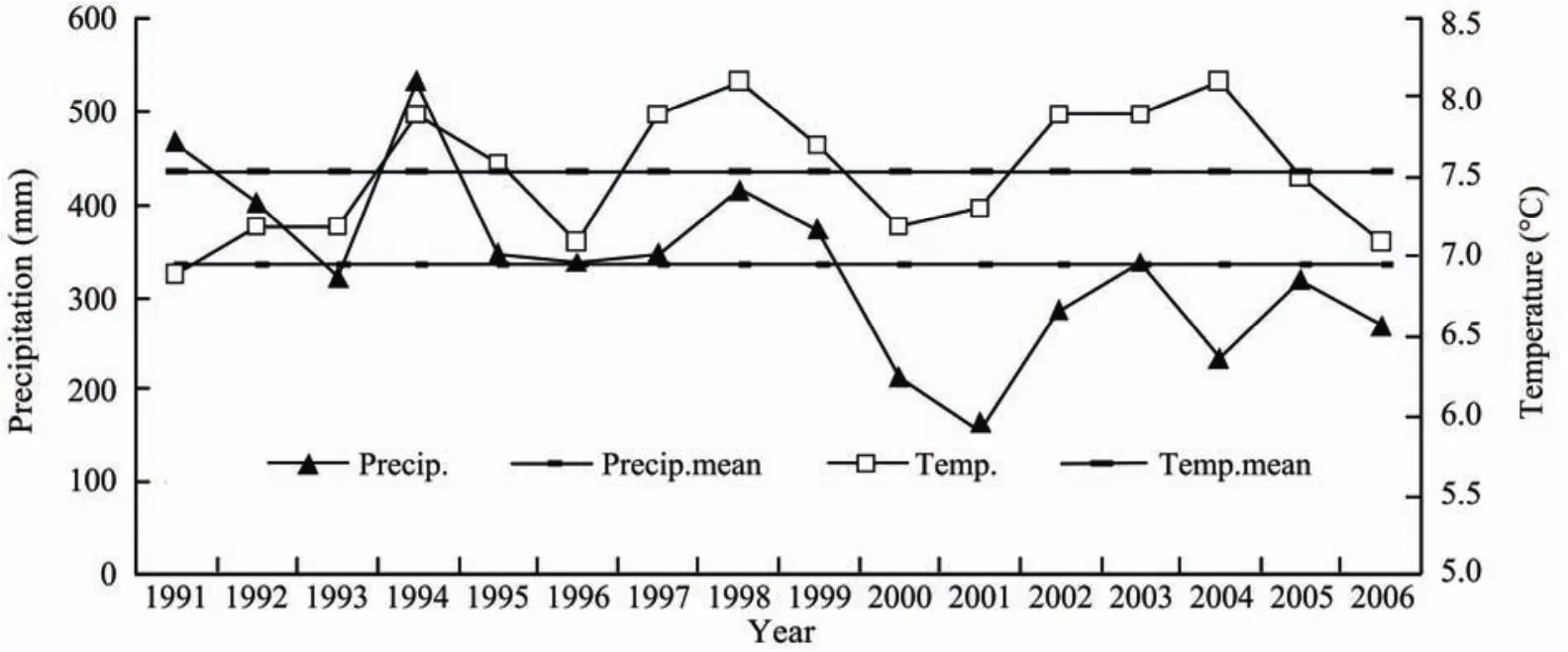
Figure 1 Changes of annual mean precipitation and air temperature during the experiment period
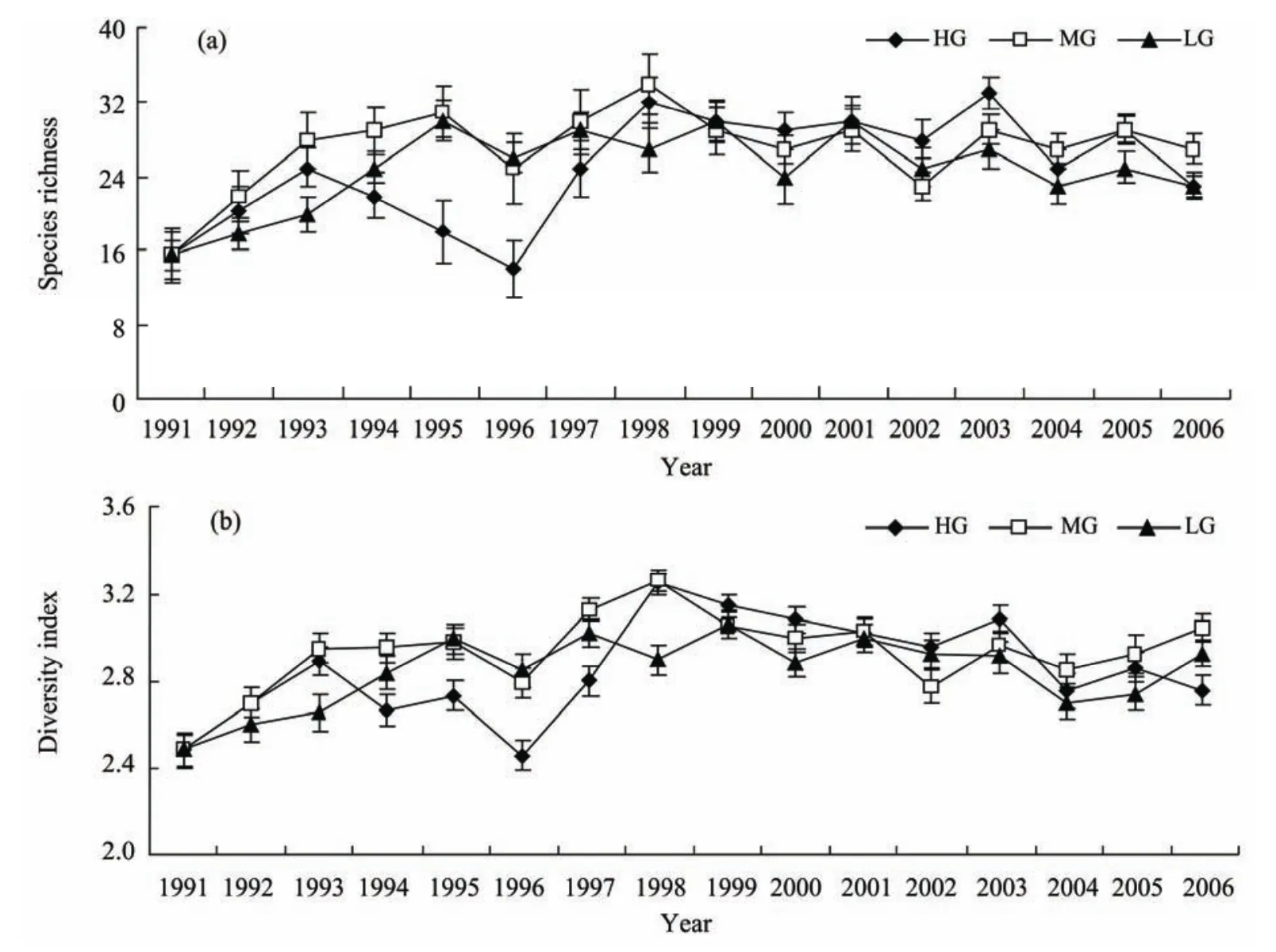
Figure 2 Changes of plant species richness (a) and diversity (b) under different grazing intensities. Bars represent means ±SD.HG = heavy grazing; MG = moderate grazing; LG = light grazing.

Figure 3 Changes in vegetation canopy coverage (a) and height (b) under different grazing intensities. Bars represent means ±SD.HG = heavy grazing; MG = moderate grazing; LG = light grazing.
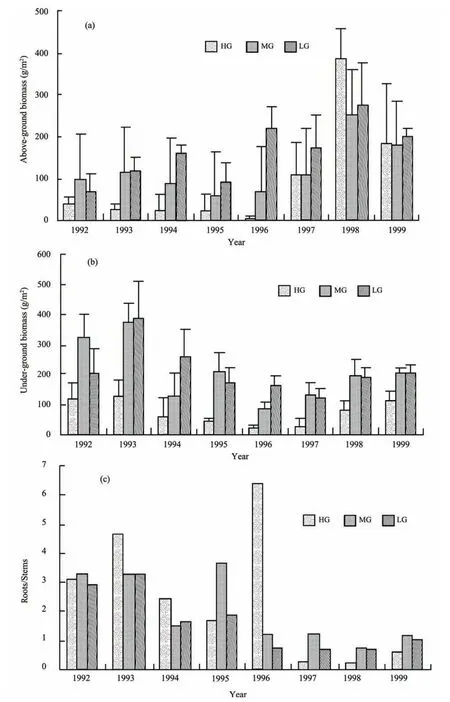
Figure 4 Changes in above-ground (a) and below-ground (b) biomass and roots/stems rates (c) under different grazing intensities.Bars represent means ±SD. HG = heavy grazing; MG = moderate grazing; LG = light grazing.
3.5. The response of tree-ring growth to precipitation
There was no significant difference in the dominant plant species of the communities among the different treatments before the grazing experiment (Table 1). The dominant species in the three treatments were altered by grazing from the second year on. The dominant species,Setaria viridisandPennisetumcentrasiaticum,were gradually replaced byChloris virgataandEragrostis pilosain the HG sites, byAristida adscensionisandLeymus secalinusin the MG sites, and byA.adscensionisandPhragmites communisin the LG sites. After the grazing was removed, the dominant species were further replaced byArtemisia scopariaandLespedeza davuricain both the HG and MG sites, but remained stable in the LG sites.
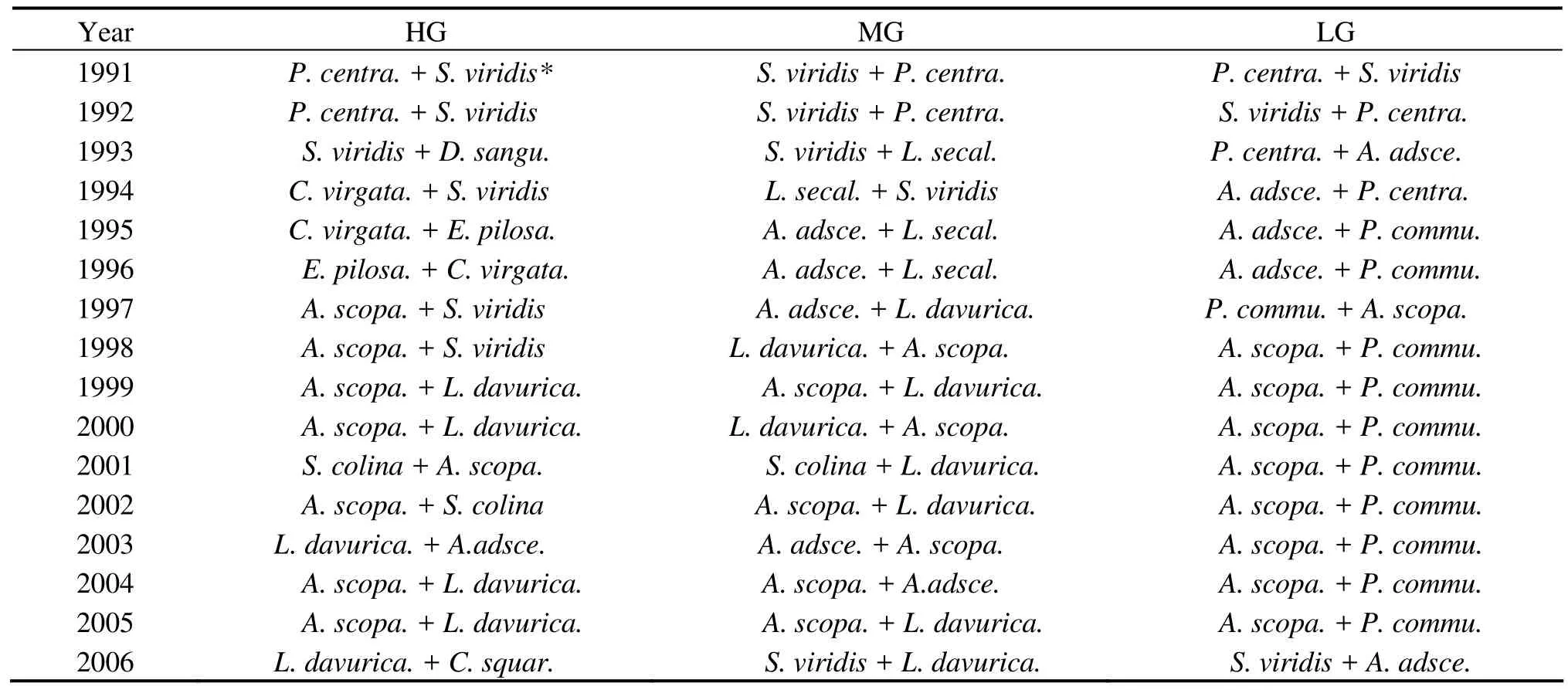
Table 1 Changes in dominant plant species of the communities during the experiment
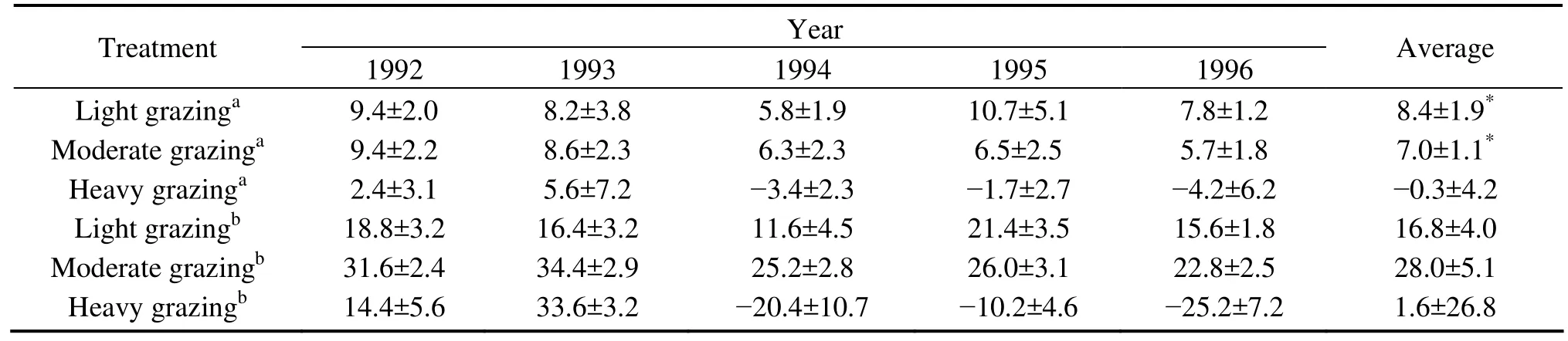
Table 2 Liveweight gain of average individual and total liveweight gain of sheep in the different treatments. Values are means ±SD.All values are significantly different (P<0.05) except those marked with *.
3.6. Sheep liveweight gain and livestock productivity
Average liveweight gain per sheep was not significantly different (P>0.05) between the LG sites and the MG sites, but at both sites the gain was significantly higher than in the HG sites during the grazing experiment period(Table 2). Average liveweight gain in the HG sites was positive in the first two years and became negative from the third year on. Net livestock productivity in the MG and LG sites was stable over the study period, and total productivity in the MG sites (140.0 kg/ha) was significant higher than that in the LG sites (83.8 kg/ha) in a 5-year experiment period. Accumulative liveweight gain was -7.8 kg/ha during the experiment period and -25.2 kg/ha in the final year of the experiment in the HG sites. The net livestock productivity in all the treatments was zero during the restoration experiment because there was no grazing during that period.
3.7. Correlation between ecosystem change and environment factors
To understand the relation of ecosystem changes with grazing intensity and climate change, we correlated the community properties and livestock productivity with grazing intensity as well as precipitation and temperature(Table 3). The results showed that in the grazing experiment, all of the community properties had a significant negative correlation with the grazing intensity (P<0.05),and no significant correlation with precipitation and temperature (P>0.05). In the restoration experiment,most of the community properties had a positive correlation with the precipitation and a negative correlation with the temperature, but no correlation coefficients reached a significant level (P>0.05) except the correlation of above-ground biomass with the precipitation (P<0.05).The sheep gain and the livestock productivity were negatively correlated with grazing intensity as well as precipitation and temperature, but the correlation was significant only between sheep liveweight gain and grazing intensity (P<0.05).
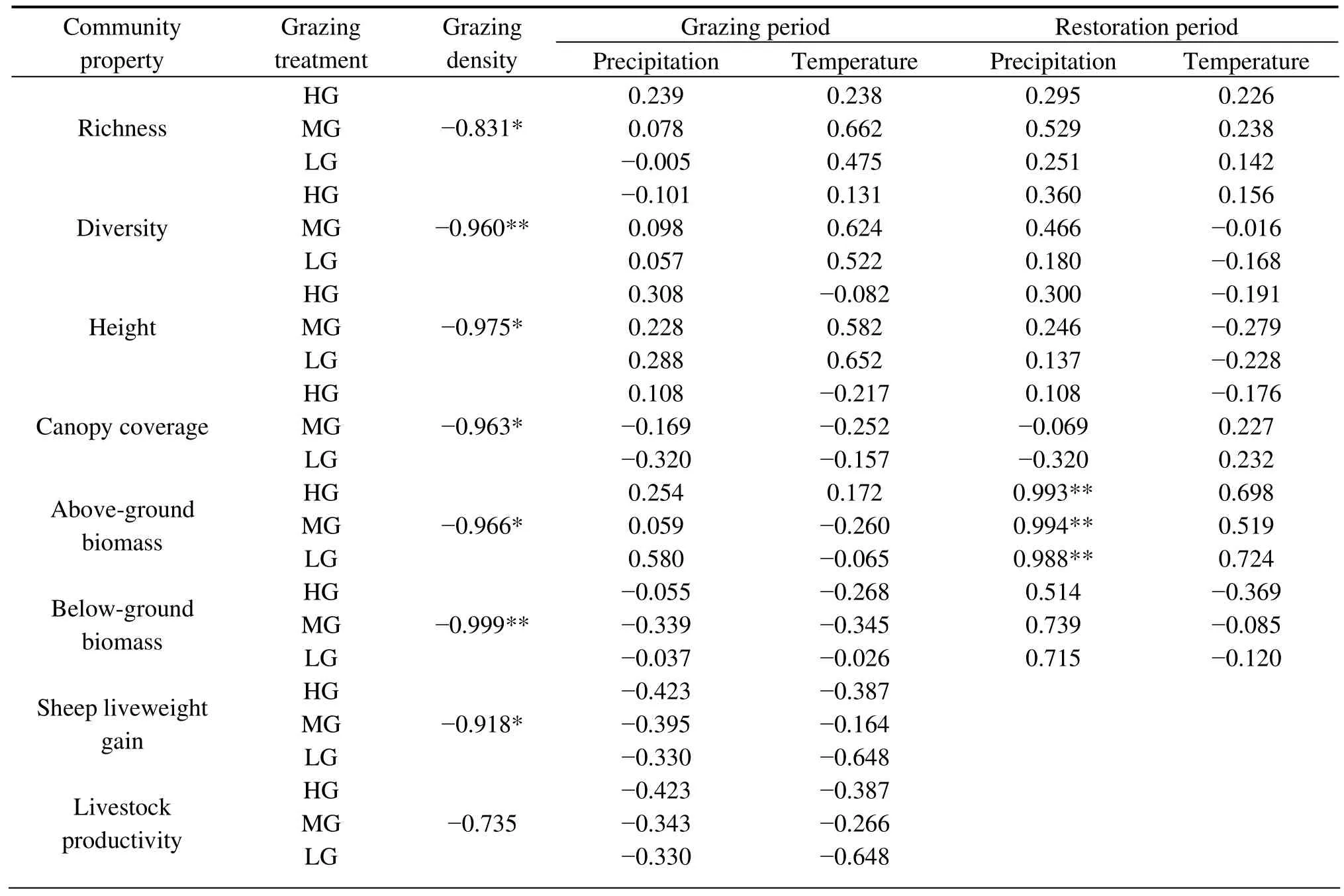
Table 3 Correlation coefficients of the vegetation index to grazing and climate changes
4. Discussion
4.1. Effects of grazing and climate change on the grassland ecosystem
Many studies have shown that livestock grazing has significant effects on grassland ecosystems (Carmel and Kadmon, 1999; Liuet al., 2002), and that the structure and function of a grassland ecosystem can be altered with changed grazing intensity (Jutila, 1999; Donget al., 2002). Some studies suggest that: (1) Sustained overgrazing can result in significant decrease of the canopy cover, height, biomass,and species richness and diversity (Donget al., 2002;Anderson and Hoffman, 2007); (2) Moderate grazing can maintain a low canopy coverage, height, and biomass, and allows a high species richness and diversity to develop(Cooperet al., 2005; Liu and Li, 2006); (3) Light grazing can promote significant increase of the plant species diversity, canopy coverage, and biomass. However, other studies indicate that the species richness and diversity are not significantly affected by sustained heavy grazing (Anderson and Hoffman, 2007), though the highest species diversity is found under moderate grazing (Zhang, 1998).
In the present study, the canopy coverage, height, and above- and below-ground biomass decreased significantly with increased grazing intensity, whereas the species rich-ness and diversity was the highest in the moderate grazing sites and lowest in the HG sites. Correlation analysis showed that the species richness, diversity, canopy coverage, height,and biomass all had a significant negative correlation with the grazing intensity, and the correlation between them and climate factors was not significant in the grazing experiment.This suggested that, on the one hand, grazing intensity was primarily responsible for the changing vegetation, and changing climate was not a restrictive factor on vegetation developing in the grazing experiment (Schultz and Halpert,1995; Thomaset al., 2003); on the other hand, greater grazing intensity had greater effects on the canopy coverage,height and biomass, but moderate grazing could maintain higher species richness and diversity than either heavy grazing or light grazing. This is consistent with the results by Bullocket al. (1994) and Noy-Meir (1998) but is not in agreement with a study by Anderson and Hoffman (2007).In addition, our results also showed that sustained heavy grazing resulted in severe disservice to grassland vegetation; moderate grazing did not cause severe degradation of the grassland vegetation (though the canopy coverage,height, and biomass were lower in the moderate grazing sites than in the light grazing sites); and light grazing can promote comprehensive restoration of degraded grassland.These results were consistent with Wanget al.(1996) and Liuet al.(2002).
Maintaining the stability of grassland vegetation and improving livestock productivity are the main aims of the grassland manager (Hooper and Vitousek, 1997; Liuet al.,2002). If only protection of vegetation was considered,light grazing or zero-grazing would be the best choice(Wanget al., 1996; Zhaoet al., 2004). But our results showed that the livestock productivity was 40% lower in the LG sites than in the MG sites; furthermore, moderate grazing did not significantly damage the grassland vegetation.This suggested that from the point of view on animal husbandry, moderate grazing should be selected for the management practice of sandy pastures in Inner Mongolia (Zhaoet al., 2003).
4.2. Effects of enclosure and climate change on the grassland vegetation
Enclosure by fence is a primary method of promoting natural restoration of degraded grassland in China (Zhaoet al., 2003). Results of previous studies suggested that cessation of grazing may lead to changes in the phenology of species and an increase in canopy cover, height, and above-ground biomass as well as species richness and diversity (Carmel and Kadmon, 1999; Liuet al., 2002). However,other studies indicated that long-term protection from grazing had been found to lead to a decline in species richness and diversity and standing biomass (Zhang, 1998; Donget al., 2002). In the present study, the species richness, diversity, canopy coverage, height, and above-ground biomass all increased by a large margin in the early period of fence enclosure; furthermore, the more severe original grazing disturbance would lead to the faster restoration rate. For example, in 1999 when grazing ceased, the canopy coverage increased by 1910.0%, 45.8%, and 5.2% compared to 1996 in the HG, MG, and LG sites, respectively. In addition, there were significant differences in the restoration rate in different properties of the vegetation. For example, in the HG sites,the above-ground biomass increased by 5460.0% and the species diversity only increased by 110.0% in 1999 compared to 1996. The results showed that after the grazing disturbance was removed, the natural restoration process of degraded vegetation was very rapid in the semi-arid sandy grassland of Inner Mongolia, and the restoration rate was faster in the canopy coverage, height, and above-ground biomass than in the richness and diversity. This is consistent with Zhang (1998) and Zhenget al.(2005).
Interestingly, the species richness, diversity, canopy coverage and height all decreased significantly in the three types of treatment areas since 2000. Generally, the order of decreased range was HG sites > LG sites > MG sites, and canopy coverage > height > richness > diversity. Apparently,this kind of vegetation change was closely related to climate changing because the early period (1996–1999) of the restoration experiment was rainy (a warm-humid climate) and the later period (2000–2006) was rainless (a warm-drought climate). This showed that changing climate was primarily responsible for changes in the vegetation after grazing was removed (Liet al., 2006; Geet al., 2007). A warm-humid climate is favorable to restoration of degraded grassland, and a warm-drought climate may result in vegetation degradation (Nautiyalet al., 2004). But correlation analysis showed that none of the correlation coefficients among the community properties and the climate factors reach a significant level (P>0.05), except for the correlation of above-ground biomass with precipitation. Zhang (1998) and Liet al.(2006)indicated that the restoration processes of grassland vegetation may be controlled by many natural factors such as climate, soil, ground water, and the vegetation itself after the grazing disturbance is removed. Our results also showed that the vegetation changes may not be controlled solely by climate change (Nautiyalet al., 2004; Liet al., 2006). What factors are actually responsible for vegetation changes needs to be further studied in the future.
4.3. Effects of grazing and climate change on community succession
In grassland ecology, succession (known as a directional change in grassland vegetation) has been discussed mainly in relation to disturbance (Zhang, 1998). However, directed change in vegetation also follows from lasting changes in environmental constraints, such as in grazing pressure and climate change (Hooper and Vitousek, 1997; Zhaoet al.,2003). Livestock grazing and climate change can influence the status of dominant plant species in the community(Carmel and Kadmon, 1999), and can even result in subro-gation of the dominant species or community succession(Cooperet al., 2005), depending on whether or not the dominant species are palatable as well as drought-resistant(Nautiyalet al., 2004).
In the present study, the dominant plant species in the three treatments were the same before the grazing experiment and in the first year. Whereas from the second year on, the original dominant species,Pennisetum centrasiaticumandSetaria viridis, were gradually displaced byEragrostis pilosaandChloris virgatain the HG sites, byAristida adscensionisandLeymus secalinusin the MG sites, and byA. adscensionisandPhragmites communisin the LG sites. This showed that there are significant differences in succession direction of the vegetation under different grazing intensities.S. viridis,E. pilosa,C. virgata,andA. adscensionisare all annual grasses; the palatability was significantly lower inE. pilosaandC. virgatathan inA. adscensionis, but their grazing resistance was just the opposite.P. centrasiaticum,L. secalinus, andP. communisare all perennial grasses, butP. centrasiaticumis a drought-resistant cluster-type grass,L. secalinusis a drought-resistant root-stem grass, andP. communisis a broadly adapted root-stem grass. These results suggest that the heavy grazing initiated a retrogressive succession of grassland vegetation, resulting in replacing a palatable perennial grass by an unpalatable annual grass. The moderate and light grazing initiated a progressive succession of grassland vegetation, resulting in replacing a cluster-type grass by a root-stem grass (Carmel and Kadmon, 1999;Zhaoet al., 2003). Jutila (1999) and Thomas (2003) indicated that grazing by livestock can alter the competitive balance between annuals and perennials. At low grazing intensity, perennials can dominate grasses through competition for light and water, but as grazing increases, the palatable annual species are reduced, allowing opportunities for grazing-resistant and unpalatable grasses to invade and increase. Our results are in agreement with their viewpoint.
After grazing disturbance was removed, the original dominant species was rapidly replaced by the annual plantArtemisia scopariaand the perennial leguminous speciesLespedeza davuricain both the HG and MG sites. The dominant species,P. communis, was not replaced by other species at all times, thoughA. adscensioniswas rapidly replaced byA. scopariain the LG sites. This showed that removal of grazing disturbance can lead to progressive succession of community, but the succession direction was different because of the original status of the disturbed vegetation (Zhang, 1998; Donget al., 2002). Generally, the vegetation succession was relatively faster in the grassland originally disturbed by heavy or moderate grazing, whereas the vegetation originally disturbed by light grazing was relatively stable. Some studies indicated that climate changes,particularly changes of precipitation and air temperature,usually result in vegetation fluctuation (Zhang, 1998; Nautiyalet al., 2004). However, our study showed that after the grazing disturbance was removed, community succession was not significantly influenced by climate change, although the species diversity, canopy coverage, and height presented fluctuating changes with changing climate. This agreed with the results by Wang (1996) and Donget al.(2002).
5. Conclusions
Our results showed that: (1) Grazing was primarily responsible for vegetation changes in grazing grassland, and the vegetation dynamic was controlled significantly by precipitation after grazing was removed. (2) Although the effects of grazing on grassland vegetation increased significantly with increase of grazing intensity, only sustained heavy grazing resulted in serious degradation of the vegetation. Moderate grazing can maintain vegetation stabilization,and light grazing can promote rapid restoration of degraded vegetation. (3) Livestock productivity was highest in the moderately grazed grassland, and consistent heavy grazing can lead to rapid decrease of livestock productivity. (4) After removing of grazing, the restoration rate of vegetation depended on the original state of the disturbed vegetation.Generally, the more severe of the original grazing disturbance leads to the faster restoration of vegetation after removal of grazing. (5) A warm-humid climate is favorable to restoration of degraded vegetation, and a sustained warm-drought climate may result in degradation of grassland vegetation. (6) Grazing pressure and climate change had greater effects on the canopy coverage, height, and biomass than on the species richness and diversity of the vegetation. (7) Heavy grazing can cause a retrogressive succession of grassland vegetation, whereas moderate and light grazing may promote progressive succession of community,and the effects of changing climate on succession processes were not significant in the short term. (8) The management practice of heavy grazing should be stopped for the sustainable use of grassland; the proper grazing intensity for the studied sandy grassland is two to three sheep or sheep equivalents per hectare in Inner Mongolia.
The authors are grateful to the anonymous reviewers for their critical review and comments on drafts of this manuscript. This research was funded by a Chinese National Key Project for Basic Scientific Research (2009CB421303) and a Chinese National Fund Project (30972422).
Anderson PML, Hoffman MT, 2007. The impacts of sustained heavy grazing on plant diversity and composition in lowland and upland habitats across the Kamiesberg Mountain range in the Succulent Karoo, South Africa. J. Arid Environ., 70: 686–700.
Augustine DJ, McNaughton SJ, 2006. Interactive effects of ungulate herbivores, soil fertility, and variable rainfall on ecosystem processes in a semi-arid savanna. Ecosystems, 9: 1242–1256.
Bullock JM, Hill BC, Dale MP, Silvertown J, 1994. An experimental study of the effects of sheep grazing on vegetation change in a species-poor grassland and the role of seedling recruitment into gaps. J. Appl. Ecol.,31: 493–507.
Carmel Y, Kadmon R, 1999. Effects of grazing and topography on long-term vegetation changes in a Mediterranean ecosystem in Israel.Plant Ecol., 145: 243–254.
Cooper A, McCann T, Ballard E, 2005. The effects of livestock grazing and recreation on Irish machair grassland vegetation. Plant Ecol., 181:255–267.
Dong QM, Li QY, Ma YS, Li FJ, Li YF, 2002. Effect of grazing intensity on above-ground biomass and vegetation structure in summer alpine meadow. Qinghai Pratacult., 11(2): 8–11.
Ge QS, Wang F, Chen PQ, Tian YU, Cheng BB, 2007. Review on global change research. A. Earth Sci., 22(4): 417–427.
Giourga H, Margaris NS, Vokou D, 1998. Effects of grazing pressure on succession process and productivity of old fields on Mediterranean islands. Env. Manag., 22: 589–596.
Hooper DU, Vitousek PM, 1997. The effects of plant composition and diversity on ecosystem processes. Science, 277: 1302–1305.
Jutila H, 1999. Effect of grazing on the vegetation of shore meadows along the Bothnian Sea, Finland. Plant Ecol., 140: 77–88.
Li X, Li XB, Wang H, Yu F, Yu HJ, Yang H, 2006. Impact of climate change on temperate grassland in northern China. J. Beijing Norm. Univ.,42(6): 618–624.
Liu Y, Wang DL, Wang X, Ba L, Sun W, 2002. The effect of grazing intensity on vegetation characteristics inLeymus chinensisgrassland. Acta Pratacult. Sin., 11(2): 22–28.
Liu ZG, Li ZQ, 2006. Plant biodiversity ofAretemisia frigidacommunities on degraded grasslands under different grazing intensities after thirteen-year enclosure. Acta Ecol. Sin., 26 (2): 475–482.
Nautiyal MC, Nautiyal BP, Vinay P, 2004. Effect of grazing and climatic changes on alpine vegetation of Tungnath, Garhwal Himalaya, India.The Environmentalist, (24): 125–134.
Noy-Meir I, Gutman M, Kaplan Y, 1989. Responses of Mediterranean grassland plants to grazing and protection. J. Ecol., 77: 290–310.
Schultz PA, Halpert MS, 1995. Global analysis of the relationships among a vegetation index, precipitation and land-surface temperature. Int. J. Remote Sens., 16: 2755–2777.
Sokal RR, Rohlf FJ, 1995. Biometry. W. H. Freeman, New York.
Thomas HA, Gregory PA, Mark EM, 2003. Changes in vegetation structure after long-term grazing in pinyon-juniper. Ecosystems, 6: 368–383.
Wang DL, Lu XI, Luo WD, 1996. Analysis to effects of different grazing density on characteristics of rangeland vegetation. Acta Pratacult. Sin.,5(3): 28–33.
Zhang W, 1998. Changes in species diversity and canopy cover in steppe vegetation in Inner Mongolia under protection from grazing. Biodivers.Conserv., 7: 1365–1381.
Zhao HL, Zhao XY, Zhang TH, Wu W, 2003. Desertification Processes and its Restoration Mechanisms in the Horqin Sand Land. China Ocean Press,Beijing.
Zhao HL, Li SG, Zhang TH, Ohkuro T, Zhou RL, 2004. Sheep gain and species diversity: in sandy grassland, Inner Mongolia. J. Range Manage.,57: 187–190.
Zhao HL, Zhao XY, Zhang TH, Zhou RL, Li SG, Ohkuro T, Drake S, 2005.Desertification processes of sandy rangeland due to overgrazing in semi-arid area, Inner Mongolia, China. J. Arid Environ., 62: 309–319.
Zheng CL, Cao ZL, Wang X, Zhao TN, Ding GD, 2005. Effects of enclosure on vegetations recovery in desertified grassland in Hulunbeir. Sci.Soil Water Conserv., 3(3): 78–81.
Zhu ZD, Chen GT, 1994. Sandy Desertification in China. Science Press,Beijing.
10.3724/SP.J.1226.2011.00223
*Correspondence to: HaLin Zhao, Prof. of Cold and Arid Regions Environmental and Engineering Research Institute, Chinese Academy of Sciences, No. 320, Donggang West Road, Lanzhou, Gansu 730000, China. Tel.: +86-931-4967201, Fax: +86-931-4967219;Email: zhaohalin9988@hotmail.com
17 January 2011 Accepted: 9 March 2011
 Sciences in Cold and Arid Regions2011年3期
Sciences in Cold and Arid Regions2011年3期
- Sciences in Cold and Arid Regions的其它文章
- Will Glacier No.1 Tianshan exist in the 22nd century?
- Sensitivity of mountain runoff to climate change for Urumqi and Kaidu rivers originating from the Tianshan Mountains
- The research of sea-land breeze (SLB) on the west coast of Bohai Bay based on field observation
- Temporal variations in aerosol composition at Lanzhou City, Northwestern China
- Hydrological and climatological glaciers observation 20 years on Tanggula Pass of Tibetan Plateau: its significance and contribution
- Concentration of trace elements and their sources in a snow pit from Yuzhu Peak, north-east Qinghai-Tibetan Plateau