
Competition, spatial pattern, and regeneration of Haloxylon ammodendron and Haloxylon persicum communities in the Gurbantunggut Desert,Northwest China
2022-10-17 09:07:36LIUYaxuanZENGYongYANGYuhuiWANGNingLIANGYuejia
Journal of Arid Land 2022年10期
LIU Yaxuan, ZENG Yong*, YANG Yuhui, WANG Ning, LIANG Yuejia
1 Xinjiang Laboratory of Lake Environment and Resources in Arid Zone, Urumqi 830054, China;
2 College of Geographic Science and Tourism, Xinjiang Normal University, Urumqi 830054, China;
3 School of Geography and Tourism, Shaanxi Normal University, Xi'an 710119, China
Abstract: Competition, spatial pattern, and regeneration are important factors affecting community composition, structure, and dynamics. In this study, we surveyed 300 quadrats from three dunes (i.e., fixed dunes, semifixed dunes, and mobile dunes) in the Gurbantunggut Desert, Northwest China, from late May to early June in 2021. The intraspecific and interspecific competition, spatial pattern, and regeneration of Haloxylon ammodendron and Haloxylon persicum were studied using the Hegyi competition index and point pattern analysis methods. The results showed that the optimal competition distance of the objective tree in the H. ammodendron and H. persicum communities was 6 m. The intraspecific and interspecific competition of H. ammodendron was the greatest in fixed dunes, while the competition intensity of H.persicum in semifixed dunes and mobile dunes was greater than that in fixed dunes. The order of competition intensity of the two populations was seedlings>saplings>adults, and the competition intensity gradually decreased with the increase in plant diameter. The spatial distribution pattern of the three life stages of H. ammodendron and H. persicum was random, and there were no correlations between seedlings and saplings, adults and saplings, and seedlings and adults. The density of regenerated seedlings and saplings of H. ammodendron in the three dunes followed the order of fixed dunes>semifixed dunes>mobile dunes, and that of H. persicum in the three dunes followed the order of mobile dunes>semifixed dunes>fixed dunes. Therefore, when artificially planting H. ammodendron and H. persicum for sand control,the planting interval should be 6 m, and seedlings should be planted next to adults to minimize the competition between plants, which can promote the renewal of H. ammodendron and H. persicum and the stabilization of the ecosystem.
Keywords: competition; spatial pattern; regeneration; Haloxylon ammodendron; Haloxylon persicum; Gurbantunggut Desert
1 Introduction
Competition occurs between two or more organisms, with the purpose of maximizing the capture of limited resources (such as water, light, and nutrients) for individual species through growth and development at the expense of other species' available resources (Craine and Dybzinski, 2013;Gao et al., 2017). In plant competition, populations are affected not only by intraspecific competition caused by the density pressure of similar populations, but also by interspecific competition caused by different populations competing for environmental resources (Chen et al.,2018). The intraspecific and interspecific competition determines the dynamic changes in the plant population within the community (Chen et al., 2021), influencing the growth and development of individual trees, the plant community structure, and the process of community succession. The result of this competition is that the competing dominant individuals often suppress and hinder the disadvantaged party, thus causing differences in the growth and development of individual plants (Xiang et al., 2015). When interspecific competition is weaker than intraspecific competition, each species is more able than its competitors to limit the growth of its own population (Adler et al., 2018). In the composition of the community structure of desert vegetation, shrub species have obvious advantages, and the species composition is relatively simple (Ban et al., 2012; Niu et al., 2019). As an important layer in the community, the shrub layer connects the tree layer and the herb layer (He et al., 2011). Studying the competition relationship and dynamics of shrub plant communities is helpful to understand the stabilization mechanism and self-organization process of vegetation. In a survey of desert shrub competition, Ji et al. (2019) discovered that shrub-shrub competition interacts with soil characteristics to limit the maximum shrub cover in the northern Chihuahuan Desert. Through the research on shrub competition, Pierce et al. (2019) discovered that shrub-shrub competition will not limit the pace of shrub proliferation during late phases of grassland to shrubland state transitions. Zhu et al.(2021) investigated the intensified interspecific water competition after afforestation withRobinia pseudoacaciain a native shrubland and discovered that it was preferable to plantR. pseudoacaciain the valley shrubland to avoid intense water competition and to control soil erosion. By studying the competition between shrubs and grasses of different ages, Morcillo et al. (2022) found that there was no effect of competition between grass and shrub seedlings on germination or survival.
Since the 1960s, many scholars have used different competition index models to study the intraspecific and interspecific competition abilities of different species, such as the Lotka-Volterra competition model (Sardar et al., 2016), the improved model of the Hegyi competition index (Long et al., 2020), and the Hegyi individual tree competition index model(Chao et al., 2017). Among the models, the Hegyi individual tree competition index model can better reflect the demand and occupation of resources by individual plants, and is currently widely used. For example, Alla and Pasho (2015) found that tree growth inAbies borisii-regisforests was generally high, but efforts should be made to control the competition among trees, as it affects their growth. Zhang et al. (2016) also investigated the intraspecific and interspecific competition intensities of the rare and endangered treeParrotia subaequalisin Wanfo Mountains Nature Reserve, Anhui Province of China, and found that interspecific competition was the main source of competitive intensity for theP. subaequalispopulation. Kang et al. (2017) studied reforestation and natural forests on the Loess Plateau in China and found that competition was the driving factor in the spatial distribution of oak forests. Xu and Liu (2018) studied the endangered plantAcer catalpifoliumin Sichuan Province of China and found thatA. catalpifoliumcompetition was mainly from interspecific sources. Xu et al. (2020) investigated the competitive intensity of the endangered plant speciesToona ciliateandTaxus chinensisin China's middle subtropical zone and found that the competitive intensity ofT. ciliateswas dominated by intraspecific competition, while that ofT. chinensiswas dominated by interspecific competition.These studies provide a theoretical basis for revealing the status of plant communities and predicting the development trends of population competition.
The spatial pattern of a population refers to the spatial distribution or configuration characteristics of individuals within a population and is the result of the combined effects of the ecological characteristics of the population, interspecific relationships, and interactions between organisms and the environment (Dang et al., 2014). The research analysis of the spatial pattern of plant communities can reflect the basic characteristics of the population and reveal the underlying ecological processes (Yang et al., 2022). Spatial distribution pattern of populations and spatial association are two important aspects of population spatial pattern studies (Li et al.,2022). A population's spatial distribution pattern indicates the distribution of individuals in horizontal space and reflects the relationship of individuals to each other in that horizontal space(Dale, 1999), which is influenced by the combination of its own biological characteristics,interspecific relationships, habitat conditions, and their interactions (Xia et al., 2016). Spatial correlation is the basis of community formation and evolution (Perry et al., 2014), which can reflect the functional relationship of individuals of different ages restricted by natural resources(Gao et al., 2014). The spatial distribution pattern analysis of plant populations usually adopts the point pattern analysis method, which can not only determine the distribution state of the population but also provide related discussion on the distribution state of the population and the relationship between the populations on any scale. This method compensates for the inadequacy of traditional quadratic sampling, which cannot fully reflect the spatial distribution characteristics of a population and the interspecific relationship, and ensures that the results of the described community structure more in line with the actual situation (Wu et al., 2019). Shrubs are an important part of the evolution of the spatial pattern and spatial association of plant species and the dynamic changes in community succession in desert areas. Studying the spatial distribution patterns and spatial associations of desert shrubs not only helps to understand the distribution characteristics and ecological functions of different species but also provides a scientific basis for the formulation of reasonable desert vegetation protection and development measures. Tirado et al. (2015) studied shrubs in an arid ecosystem and discovered thatWhitania frutescensandMaytenus senegalensis, two shrub species that closely interact in their community, benefit from their spatial relationship. Zhang et al. (2021) investigated the spatial pattern and spatial association of shrub species in a desert dominated by relictHelianthemum songaricumand discovered that the entire population ofH. songaricumand individuals of various size classes were distributed randomly and regularly at most spatial scales in a gravel habitat but tended to form a clumped distribution in a rocky habitat. Zhang et al. (2022) discovered that two desert shrubs showed positive interspecific relationships on a small scale and negative relationships as interspecific competition intensified in the presence of grazing disturbance by studying the spatial distribution pattern and interspecific relationship of the two desert shrubs. The spatial pattern of plants depends on factors such as internal plant characteristics, interspecific relationships, and the interaction between different individuals of the same species, as well as the habitat conditions of the species and the spatial dynamics of growing competition between plants (Wang, 2019).Therefore, analyzing the competition index and spatial distribution pattern of the main desert tree species to understand the species population in the current environment, the living status of the community, and the future development trend is more helpful to protect the desert community.
The natural regeneration of plants is the way for the ecosystem to accomplish self-reproduction and restoration, and it is also the basis for maintaining the dynamic stability and sustainable utilization of the community. Natural regeneration is considered to be the gradual restoration of the structure, function, and composition of the pre-disturbed ecosystem (Chazdon and Guariguata,2016). The natural regeneration process of plants is affected by soil factors (Wang et al., 2017),stand factors (Zhu, 2021), stand age (Zhang et al., 2015), habitat (Hu et al., 2015), and other factors. Tree age determines the number of reproducing individuals and the long-term development of the plant ecosystem (Sharma et al., 2016). Regeneration requires an adequate number of seedlings and saplings; conversely, an inadequate number indicates a poor regeneration capacity (Ali et al., 2019). The regeneration of seedlings and saplings is an important stage in the natural regeneration of vegetation, and it is also a period in the life history of plants when the individual growth is fragile and sensitive to environmental changes, and the mortality rate is high;it has an important impact on the stability of the population and the succession of the community(D'Amato et al., 2009).
The Gurbantunggut Desert is located in the middle of the Junggar Basin in Xinjiang Uygur Autonomous Region of China; it is a temperate desert with rich plant diversity and is the second largest desert in China. Due to human activities and natural disturbances, desert habitats have been destroyed, local vegetation has degraded significantly, species diversity has been lost, and ecosystem functions have been severely degraded, causing serious losses to the surrounding environment and sustainable economic development.Haloxylon ammodendronandHaloxylon persicumare the main constructive species in the Gurbantunggut Desert, both belong to the family Chenopodiaceae; their leaves degenerate into scales and have strong resistance to drought and salinity. They play important roles in maintaining the regional ecological balance and promoting regional sustainable development and are listed as national third-level endangered protected plants (Dai et al., 2013). At present, research onH. ammodendronandH. persicummainly focuses on photosynthesis (Hu et al., 2021), drought resistance (He et al., 2021),population structure (Song et al., 2021), plant diversity (Zeng et al., 2016), seed germination(Tobe et al., 2000), and biomass (Xu et al., 2017). However, there is little research on the competition, spatial pattern, and regeneration ofH. ammodendronandH. persicum.
2 Materials and methods
2.1 Study area
The study area (44°10′-46°19′N, 84°31′-90°00′E) is approximately 5.11×104km2and is located in the Gurbantunggut Desert in Xinjiang Uygur Autonomous Region of China; the dune types are mainly fixed dunes, semifixed dunes, and mobile dunes, of which fixed dunes and semifixed dunes account for 87% of the total desert area, and the orientation of the dunes varies roughly from northwest to southeast, with a height of 15-20 m (Li et al., 2021). The natural vegetation in the study area is dominated by the small trees (H. ammodendronandH. persicumas the dominant species), low semi-shrubs are dominated byEphedra distachya, and herbaceous plants are dominated byErodium oxyrrhynchum,Hyalea pulchella,Ceratocarpus arenarius, andSalsola ruthenica(Zhang et al., 2022).
2.2 Research methods
2.2.1 Data collection In 2021, a total of three sites (100 m×100 m) with good protection, good growth, and a large distribution area ofH. ammodendronandH. persicumwere established in fixed dunes, semifixed dunes, and mobile dunes of the Gurbantunggut Desert. The geographical coordinates of the three sites are 44°22′N, 87°52′E; 44°36′N, 88°14′E; and 44°56′N, 88°33′E, respectively. The sites include central dunes, lower dunes, upper dunes, and tops of dunes. Each site was divided into 100 quadrats (10 m×10 m) using the adjacent grid method, with a total of 300 quadrats. Vegetation was surveyed from late May to early June. Species name, number, density, height, crown width, basal diameter, and coordinates were recorded in each quadrat. In order to determine the precipitation characteristics of the sites, we analyzed the meteorological data. However, no weather stations are located near the sites. The forecast precipitation data from European Centre for Medium-Range Weather Forecasts were used. Focusing on a grid of 0.125° latitude and longitude in 2015 and 2019, we calculated the average annual precipitation and annual average temperature. Previous research has shown that this method is highly accurate (Liu et al., 2018). In each site, five soil samples at a depth of 0-20 cm were randomly collected and mixed. The soil samples were air-dried and passed through a 2-mm sieve. Soil conductivity was determined by leaching using air-dried soil and distilled water (5:1 water:soil ratio) and was measured when the leaching solution was 25°C. The dry residue method was used to determine total salt. The aluminum box drying method was used to determine the soil water content indoors. A 0.25-mm sieve was used to filter the soil samples, and soil organic matter was determined by the external heating method of potassium dichromate. During the study, each site was located using Global Position System to record its longitude, latitude, and elevation. In each quadrat, the target trees ofH. ammodendronandH. persicumwere taken as the center of the circle to measure the distances between all the competing trees and the target trees in the sample circle. The spatial coordinate information ofH.ammodendronandH. persicumin the three sites is shown in Figure 1.
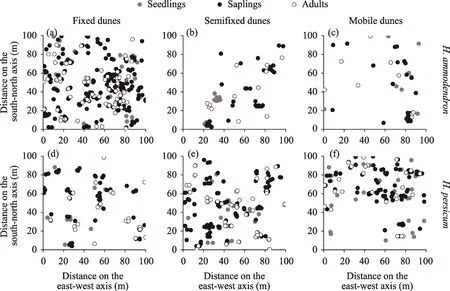
Fig. 1 Spatial distribution maps of Haloxylon ammodendron (a-c) and Haloxylon persicum (d-f) in fixed dunes,semifixed dunes, and mobile dunes
2.2.2 Determination of the optimal range of competition betweenHaloxylon ammodendronandHaloxylon persicum
A total of 106H. ammodendronand 101H. persicumwith different basal diameters were randomly selected in fixed dunes, semifixed dunes, and mobile dunes as target trees. The sample circle was set up with each target tree as the center of the circle and other trees within the sample circle as competing trees. The average competition intensity of the competing trees in circles with radii of 1-16 m (with a step difference of 1 m) ofH. ammodendronandH. persicumwas measured. The results were fitted in segments to form regression plots between different circle radii and the average competition intensity. If both fitted equations reached a significant level andR2was high, it indicated that this point was the turning point, and the radius of the sample circle corresponding to this point was the competition range of the competitive trees.
2.2.3 Classification criteria for different life stages ofH. ammodendronandH. persicum
In this study, we classifiedH. ammodendronandH. persicuminto three life stages (Jia and Lu,2004): seedlings (basal diameter<1.2 cm), saplings (1.2 cm≤basal diameter≤6.5 cm), and adults(basal diameter>6.5 cm). Hegyi competition index, Ripley'sKfunction (K(r)), and Ripley'sgfunction (g(r)) were used to analyze the competition, spatial pattern, and regeneration ofH.ammodendronandH. persicumin the three life stages.
2.3 Data analysis
2.3.1 Competition index model
The distance-related competition index model proposed by Hegyi was used to calculate the intraspecific and interspecific competition indices ofH. ammodendronandH. persicum. The equations can be expressed as follows:


whereiis the quantity of the target trees;jis the quantity of the competitive trees; CIiis the competition index of theithtarget tree; Diis the diameter at breast height of target treei(cm);Djis the diameter at breast height of competitive treej(cm);Lijis the distance between target treeiand competitive treej(m);Nis the number of competitive trees of target treei; and CI is the competition index of the species. The larger the competition index value, the more intense the competition among tree species, i.e., the greater the competitive pressure on the target tree. SinceH. ammodendronandH. persicumare shrub-like small tree plants, the basal diameter was used instead of the diameter at breast height in the single-tree competition index model (Li et al.,2016).
2.3.2 Relationship between the competition intensity and basal diameter
Regression analysis was carried out using the competition intensity and basal diameter of the target trees ofH. ammodendronandH. persicum. The competition index was the dependent variable, and the basal diameter of the target tree was the independent variable; the fitting relationship of power function regression is as follows:

whereD is the diameter at breast height of the target tree (cm); andAandBare model parameters.
2.3.3 Analysis of the spatial point pattern
Spatial point pattern ofH. ammodendronandH. persicumin different life stages was analyzed in fixed dunes, semifixed dunes, and mobile dunes. This method takes the coordinates of individual plant species in space as the basic data and conducts pattern analysis on the basis of their spatial distribution maps (Fig. 1). The multiscale spatial distribution patterns of plant species and the multiscale spatial association between the two species were studied using spatial point pattern analysis.K(r) was calculated as follows:
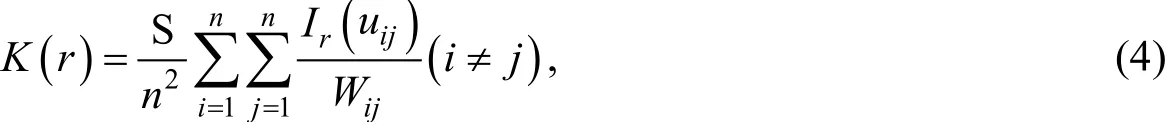
where S is the area of the study area (m2);nis the number of individuals;uijis the distance betweenipoint andjpoint;Ir(uij) is the indicator function, whenuijis less than or equal to the distance between trees,Ir(uij) is equal to 1; whenuijis greater than the distance between trees,Ir(uij) is equal to 0; andWijis the weight value used for edge correction. Theg(r) is derived fromK(r) and overcomes the cumulative effect ofK(r), which describes the probability of species occurrence within a specified circular width with any point as the center and a certain distance(scale) as the radius (Wiegandet al.,2004). Theg(r) includes the univariate correlation function ofg(r) (i.e.,g11(r)) and the bivariate correlation function of g(r)(i.e.,g12(r)). Theg11(r) was used to analyze the spatial distribution pattern of a single species, and the expression is as follows:

where dK(r) is the derivative of the functionK(r); and dris the derivative of the radiusr. Wheng11(r) is equal to 1, it indicates random distribution;g11(r)greater than 1 indicates aggregated distribution; andg11(r) less than 1 indicates uniform distribution. Theg12(r) was used to analyze the spatial association between the two species; that is, the analysis takes an individual of a species as the center, a distance (scale) as the radius, and another species within the specified circular width probability of occurrence (Zeng et al., 2019). The expression ofg12(r) is as follows:

where dK12(r) is the derivative of the functionK(r). Wheng12(r) is equal to 1, it indicates that there is no correlation between the two species;g12(r)greater than 1 indicates that the two species are positively correlated; andg12(r) less than 1 indicates that the two species are negatively correlated.
2.3.4 Population renewal analysis
The renewal density of a sample circular area is calculated as: renewal density=(number of seedlings+number of saplings)/circle area. The seedlings and saplings in the three sites were used as regeneration trees, and the adults were used as mother trees. With a single mother tree as the center, the number of surviving and dead regeneration trees within the radius of 2, 4, and 6 m sample circles were counted.
2.4 Statistical analysis
Data processing was carried out mainly through Programita, and the goodness-of-fit test was used to analyze the significance of the results. The spatial scale was 0-50 m, the circular width was set to 1 m, and the step length was 10 m. A total of 199 simulations were performed using the Monte Carlo method, and the fifth-highest and fifth-lowest simulated values were selected to construct 95% confidence intervals.
3 Results
3.1 Overview of target trees and competitive trees of H. ammodendron and H. persicum
There were 207 target trees and 618 competitive trees ofH. ammodendronandH. persicum(Table 1). ForH. ammodendron, there were 76 seedlings and saplings and 30 adults, each accounting for 71.70% and 28.30% of the total number ofH. ammodendron, respectively; forH.persicum, there were 78 seedlings and saplings and 23 adults, each accounting for 77.23% and 22.77% of the total number ofH. persicum, respectively. The number of target trees ofH.ammodendronandH. persicumshowed that the proportions of seedlings and saplings were high,and the age structure of the population tended to be young.
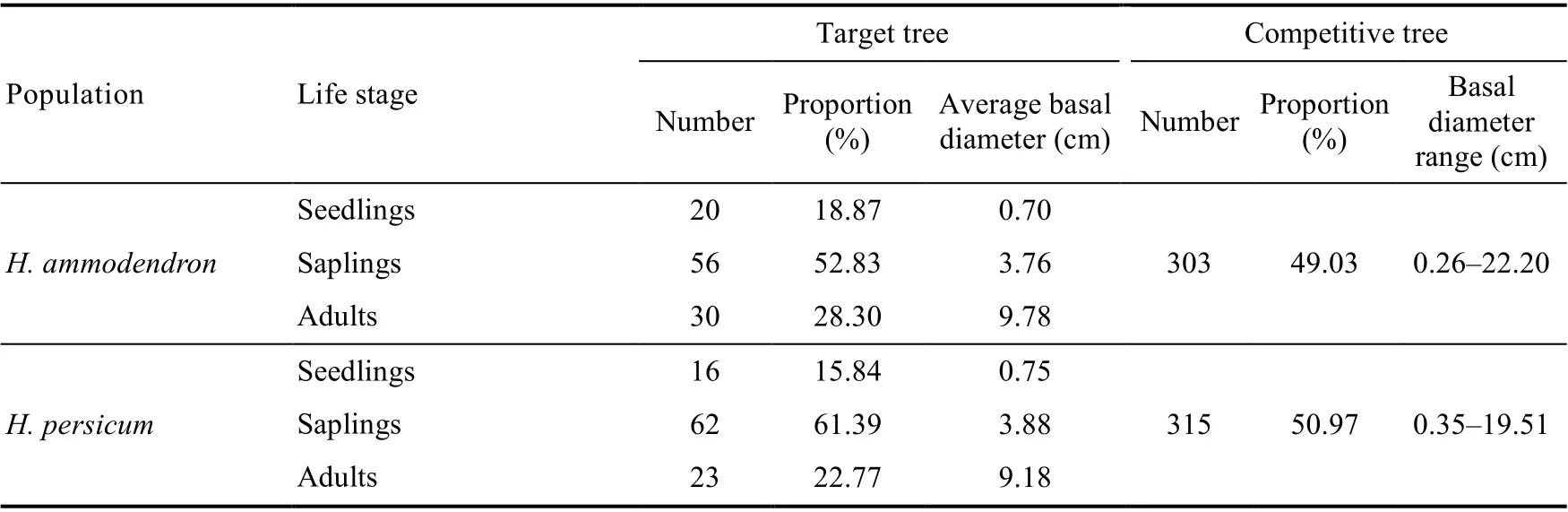
Table 1 Overview of target trees and competitive trees of Haloxylon ammodendron and Haloxylon persicum
3.2 Competition intensity of H. ammodendron and H. persicum
3.2.1 Neighborhood optimal competition range determination According to the regression relationship between the radius of the round quadrats and the average competition index ofH. ammodendronandH. persicum, the average competition index of competitive trees decreased with the increasing radius of the round quadrats, with an obvious turning point at 6 m (Fig. 2). The competition intensity ofH. ammodendronandH. persicumdecreased with the increasing radius of the round quadrats when the radius of the round quadrats was within 6 m. The fitting equation also showed that the two fitting equations ofH.ammodendronandH. persicumreached a significant level, at which point theR2was the highest.This point was the turning point, and the radius of the round quadrats corresponding to this point was the competition range of the competitive trees. Therefore, the competition range of bothH.ammodendronandH. persicumwas 6 m.

Fig. 2 Relationship of the radius of the round quadrats with the average competition intensity for H.ammodendron and H. persicum
3.2.2 Intraspecific and interspecific competition intensities betweenH. ammodendronandH.persicumin different dunes
In different dunes, with increasing stand age, the competition intensities ofH. ammodendronandH. persicumwere different. The competition intensities ofH. ammodendronandH. persicumwere the highest among the seedlings, followed by saplings and then adults (Table 2). The competition betweenH. ammodendronandH. persicumin fixed dunes, semifixed dunes, and mobile dunes was mostly intraspecific. The competition ofH. ammodendronin fixed dunes was greater than that ofH. persicum, and the competition ofH. ammodendronin semifixed dunes and mobile dunes was less than that ofH. persicum.The average intraspecific and interspecific competition intensities ofH. ammodendronfollowed the order of fixed dunes>semifixed dunes>mobile dunes. The average intraspecific competition intensity ofH. persicumwas greater in mobile dunes and semifixed dunes than in fixed dunes, and the order of average interspecific competition intensity was mobile dunes>semifixed dunes>fixed dunes.
The average intraspecific and interspecific competition intensities ofH. ammodendronseedlings, saplings, and adults followed the order of fixed dunes>semifixed dunes>mobile dunes.The average intraspecific competition intensity ofH. persicumseedlings followed the order of mobile dunes>semifixed dunes>fixed dunes, and the order of average intraspecific competition intensity of saplings and adults was semifixed dunes>mobile dunes>fixed dunes. The order of the average interspecific competition intensity ofH. persicumseedlings, saplings, and adults was mobile dunes>semifixed dunes>fixed dunes.
3.2.3 Relationship between the competition intensity and target tree basal diameter
The relationships between the basal diameter and competition index of target trees ofH.ammodendronandH. persicumwere analyzed in fixed dunes, semifixed dunes, and mobile dunes(Fig. 3). With the increase in the target tree basal diameter, the competitive intensity ofH.ammodendronandH. persicumgradually decreased. When the basal diameter of target tree exceeded 15 cm, the competition intensity tended to be stable and remained at a low level.H.ammodendronfollowed a power function, with CI=4.847D-0.726(R2=0.338,P<0.001),CI=4.369D-0.731(R2=0.711,P<0.001), and CI=3.100D-0.127(R2=0.910,P<0.001) in fixed dunes,semifixed dunes, and mobile dunes, respectively.H. persicumfollowed a power function, with CI=3.605D-0.840(R2=0.796,P<0.001), CI=5.577D-1.017(R2=0.312,P<0.001), and CI=4.374D-0.870(R2=0.463,P<0.001) in fixed dunes, semifixed dunes, and mobile dunes, respectively. Therelationship betweenH. ammodendronand the competition index was CI=4.515D-0.776(R2=0.359,P<0.001). The relationship betweenH. persicumand the competition index was CI=4.443D-0.792(R2=0.412,P<0.001). The relationship between the two populations and the competition index was CI=4.477D-0.784(R2=0.379,P<0.001).
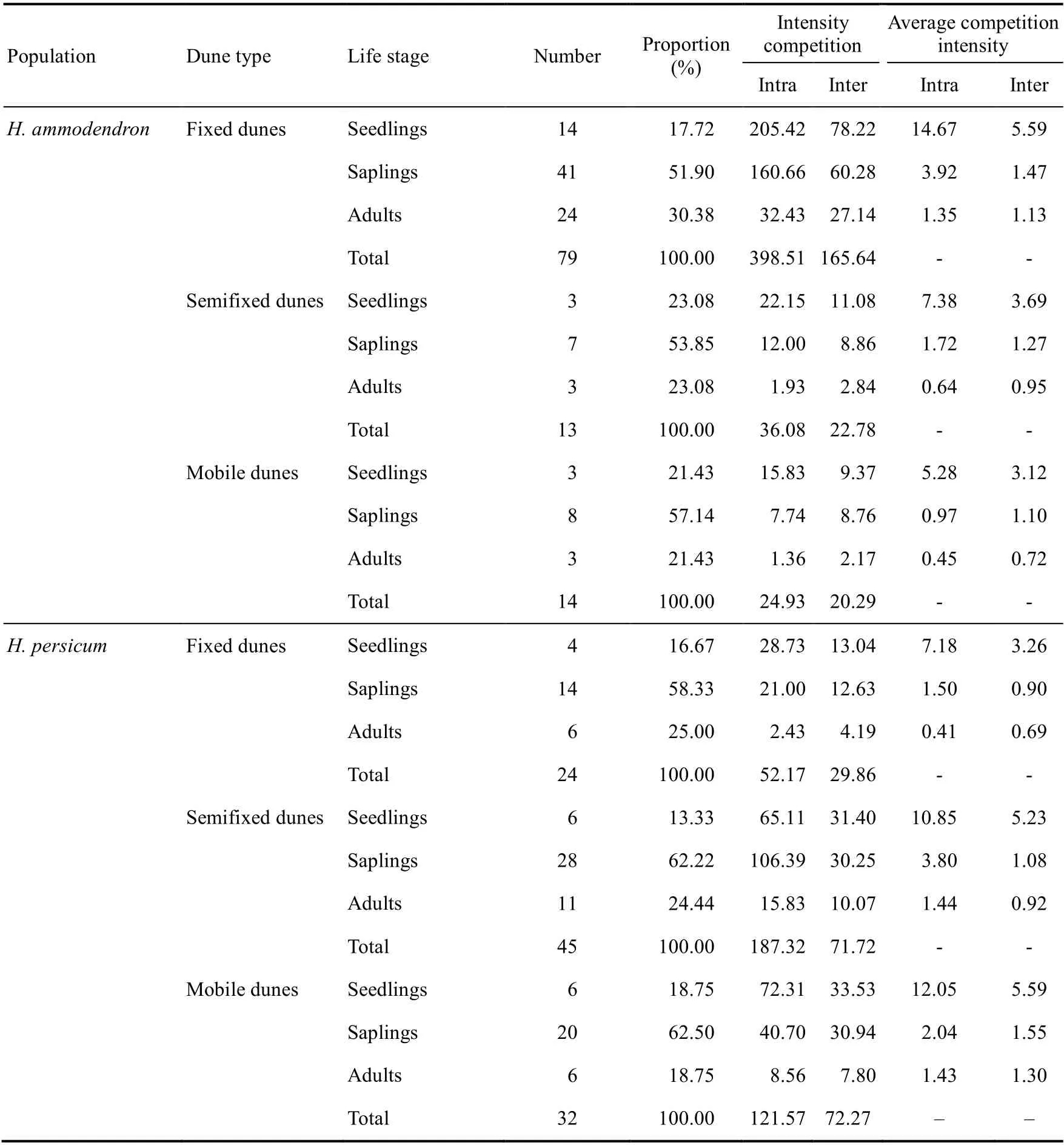
Table 2 Intraspecific and interspecific competition intensity between H. ammodendron and H. persicum
3.3 Analysis of the spatial pattern of H. ammodendron and H. persicum
3.3.1 Spatial distribution pattern ofH. ammodendronandH. persicumin different life stages
The seedlings ofH. ammodendronwere randomly distributed in fixed dunes and mobile dunes. In semifixed dunes, the distribution of the seedlings ofH. ammodendronwas mainly random but uniform at the 3, 7-9, 11-18, and 29-32 m scales and clustered at the 48 and 50 m scales (Fig. 4).The saplings ofH. ammodendronwere randomly distributed in fixed dunes. In semifixed dunes,the distribution of the saplings ofH. ammodendronwas mainly random and only clustered at the 18 and 40 m scales. In mobile dunes, the distribution of the saplings ofH. ammodendronwas mainly random and clustered at the 23 m scale. The adults ofH. ammodendronwere mainly randomly distributed in fixed dunes, clustered at the 1 m scale and uniformly distributed at the 10 and 12-13 m scales. In semifixed dunes, the distribution of the adults ofH. ammodendronwas mainly random, clustered at the 10 and 20 m scales, and uniform at the 28, 31, 36, and 48 m scales. In mobile dunes, the distribution of the adults ofH. ammodendronwas random.
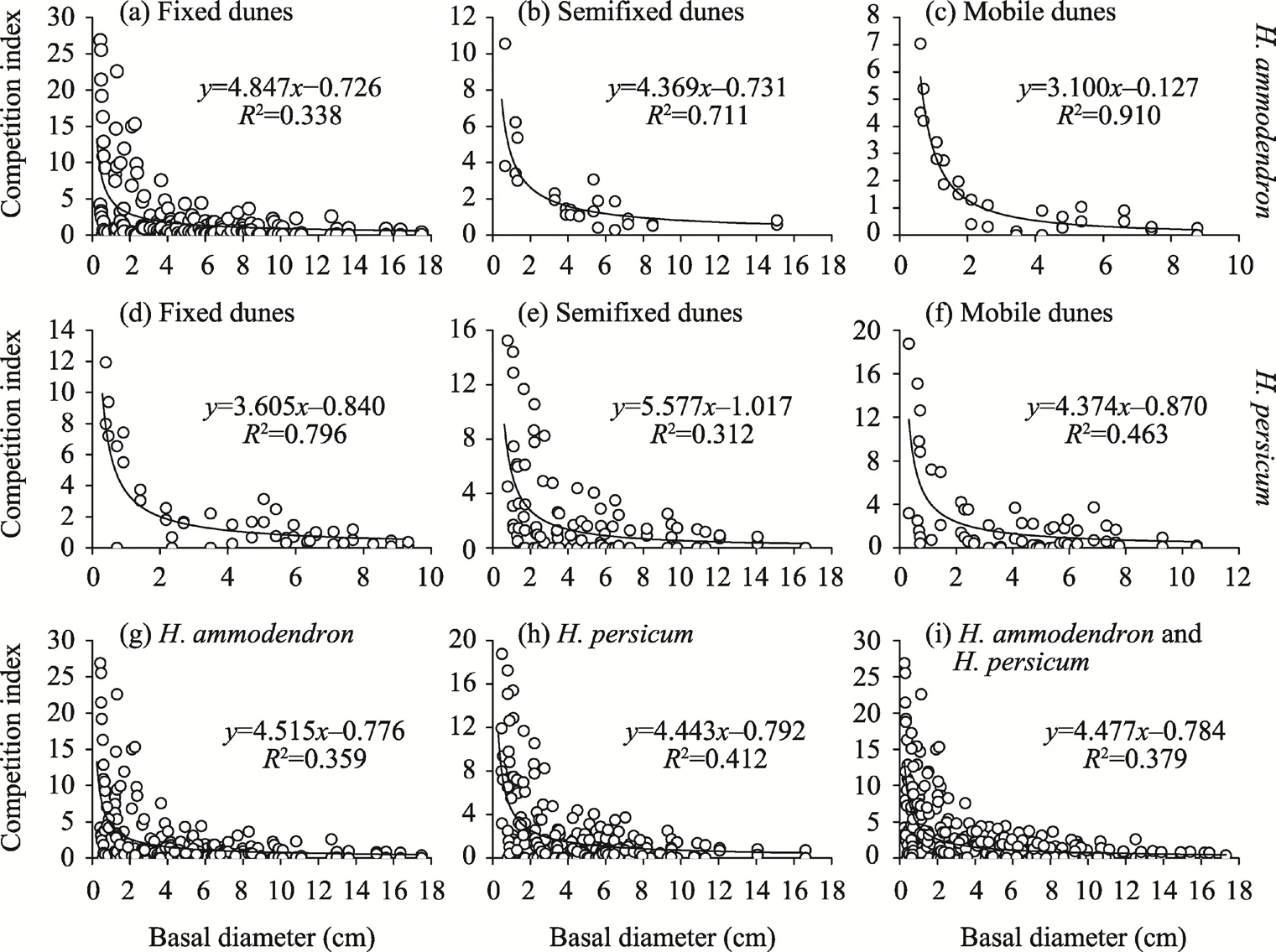
Fig. 3 Relationships of the basal diameter with competition index of target trees of H. ammodendron (a-c) and H. persicum (d-f) in fixed dunes, semifixed dunes, and mobile dunes, as well as the relationships of the basal diameter with competition index of all H. ammodendron individuals (g), all H. persicum individuals (h), and the sum of all H. ammodendron and H. persicum individuals (i).
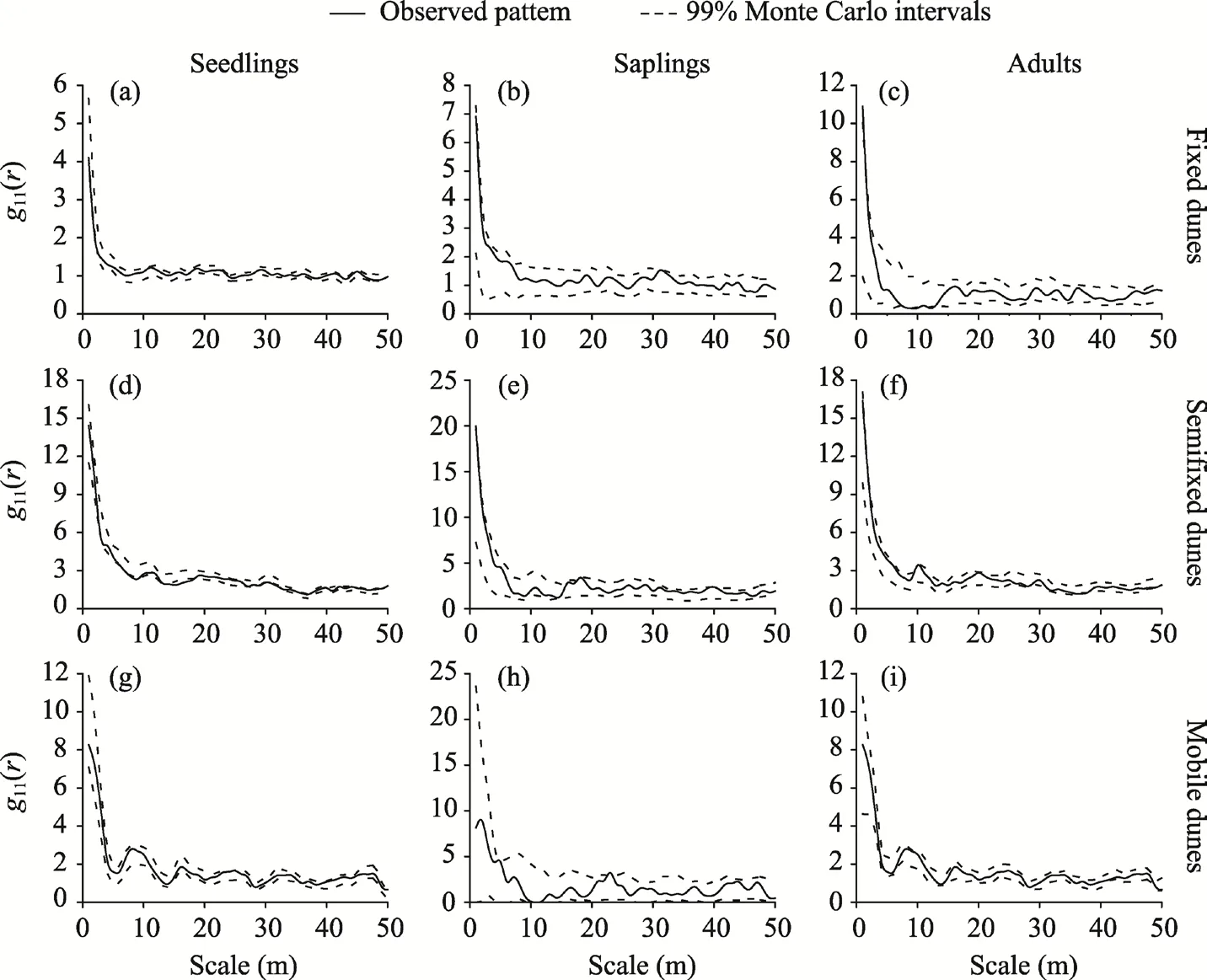
Fig. 4 Spatial distribution pattern of H. ammodendron in different life stages in fixed dunes (a-c), semifixed dunes (d-f), and mobile dunes (g-i). The g11(r) represents the spatial distribution of a population.
In fixed dunes,the seedlings ofH. persicumwere mainly randomly distributed and uniformly distributed at the 50 m scale. In semifixed dunes, the seedlings ofH. persicumwere mainly randomly distributed and aggregated at the 31 m scale. In mobile dunes, the seedlings ofH.persicumwere randomly distributed (Fig. 5). In fixed dunes, the saplings ofH. persicumwere mainly randomly distributed and clustered at the 46 and 50 m scales. In semifixed dunes, the saplings ofH. persicumwere randomly distributed. In mobile dunes, the saplings ofH. persicumwere mainly randomly distributed and clustered at the 8 and 27 m scales and uniformly distributed at the 47 m scale. In fixed dunes, the adults ofH. persicumwere mainly randomly distributed and uniformly distributed at the 45 m scale. In semifixed dunes, the adults ofH.persicumwere randomly distributed. In semifixed dunes, the adults ofH. persicumwere mainly randomly distributed. In mobile dunes, the adults ofH. persicumwere mainly randomly distributed and clustered at the 31 m scale.
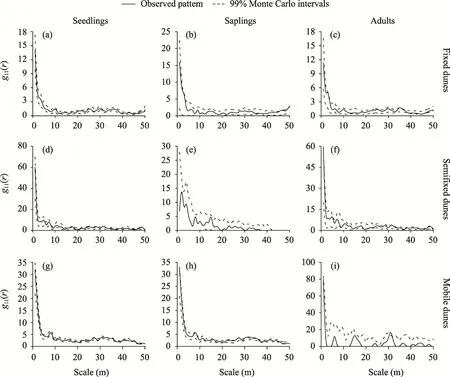
Fig. 5 Spatial distribution pattern of H. persicum in different life stages in fixed dunes (a-c), semifixed dunes(d-f), and mobile dunes (g-i)
3.3.2 Spatial association analysis ofH. ammodendronandH. persicumin different life stages
The seedlings and saplings ofH. ammodendronhad no spatial correlation in fixed dunes and mobile dunes. In semifixed dunes, the seedlings and saplings ofH. ammodendronwere mainly uncorrelated, with a significant negative correlation only at the 0-2 m scale and significant positive correlations at the 9, 11, 13-18, and 30-32 m scales (Fig. 6).The seedlings and adults ofH. ammodendronhad no spatial correlation in mobile dunes. In fixed dunes, the seedlings and adults ofH. ammodendronmainly had no spatial correlation and had a negative correlation only at the 1 m scale. In semifixed dunes, the seedlings and adultsofH. ammodendronmainly had no spatial correlation, had significant positive correlations only at the 11-15 and 43 m scales, and had a significant negative correlation at the 0-5 m scale. The saplings and adults ofH. ammodendronshowed no spatial correlation in fixed dunes. In semifixed dunes, the saplings and adultsofH.ammodendronmainly showed no spatial correlation, showed significant negative correlations only at the 0-2 and 5 m scales, and showed significant positive correlations at the 26, 28, 36-38, and 47-49 m scales. In mobile dunes, the saplings and adults ofH. ammodendronmainly showed no spatial correlation and showed a significant positive correlation only at the 12 m scale.
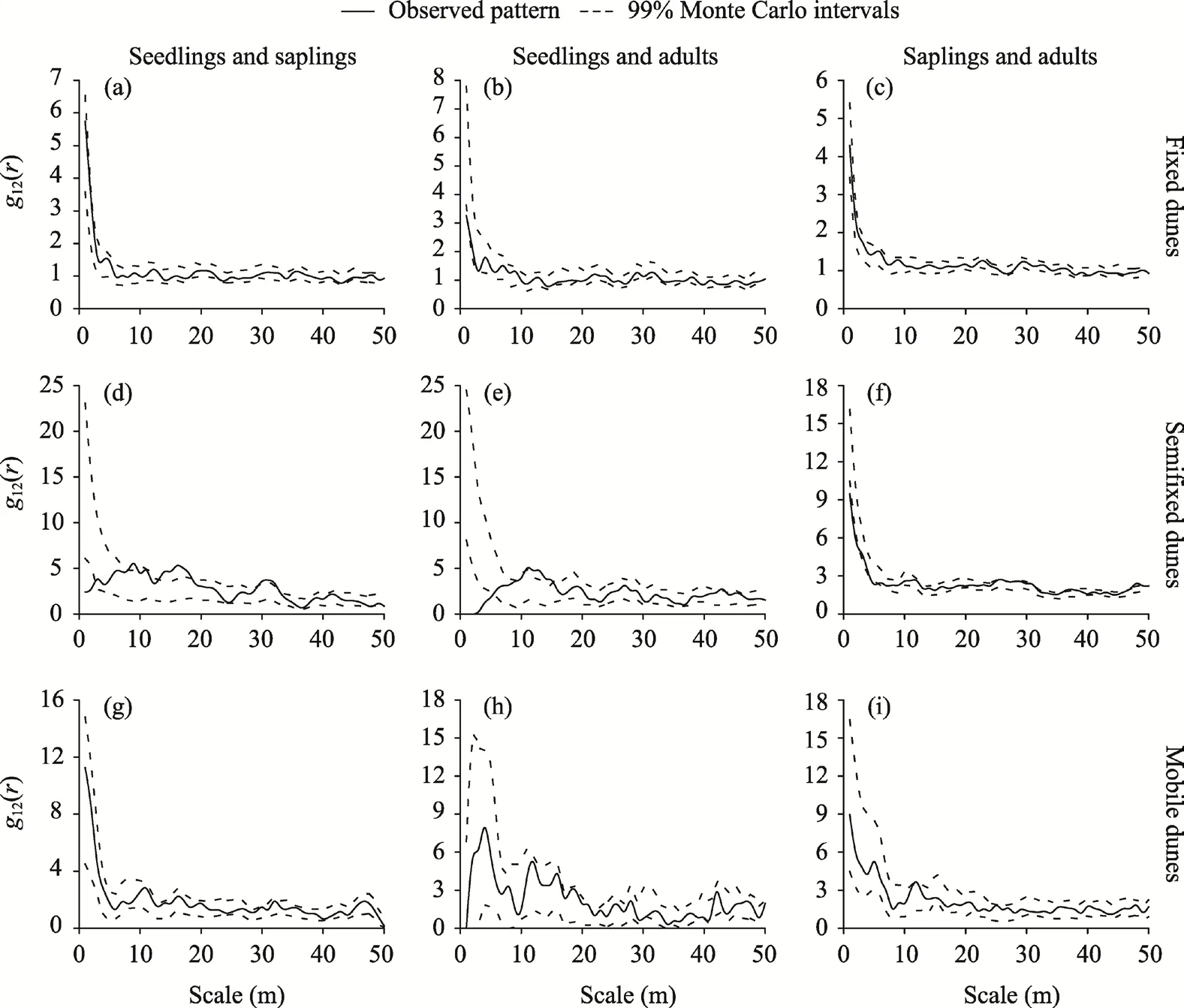
Fig. 6 Spatial association of H. ammodendron in different life stages in fixed dunes (a-c), semifixed dunes(d-f), and mobile dunes (g-i). The g12(r) represents the spatial association of the three life stages of a population.
The seedlings and saplings ofH. persicumhad no spatial correlation in fixed dunes and mobile dunes. In semifixed dunes, the seedlings and saplingsofH. persicummainly had no spatial correlation and had a significant positive correlation only at the 42 m scale (Fig. 7). The seedlings and adults ofH. persicumhad no spatial correlation in fixed dunes, semifixed dunes, and mobile dunes. The saplings and adults ofH. persicummainly had no spatial correlation and had a significant negative correlation in fixed dunes only at the 3 m scale. In semifixed dunes, the saplings and adultsofH. persicummainly had no spatial correlation, had significant negative correlations at the 1 and 23 m scales, and had significant positive correlations at the 30, 35, and 10-12 m scales. In mobile dunes, the saplings and adultsofH. persicummainly had no spatial correlation, had a negative correlation at the 27 m scale, and had a positive correlation at the 47-48 m scale.
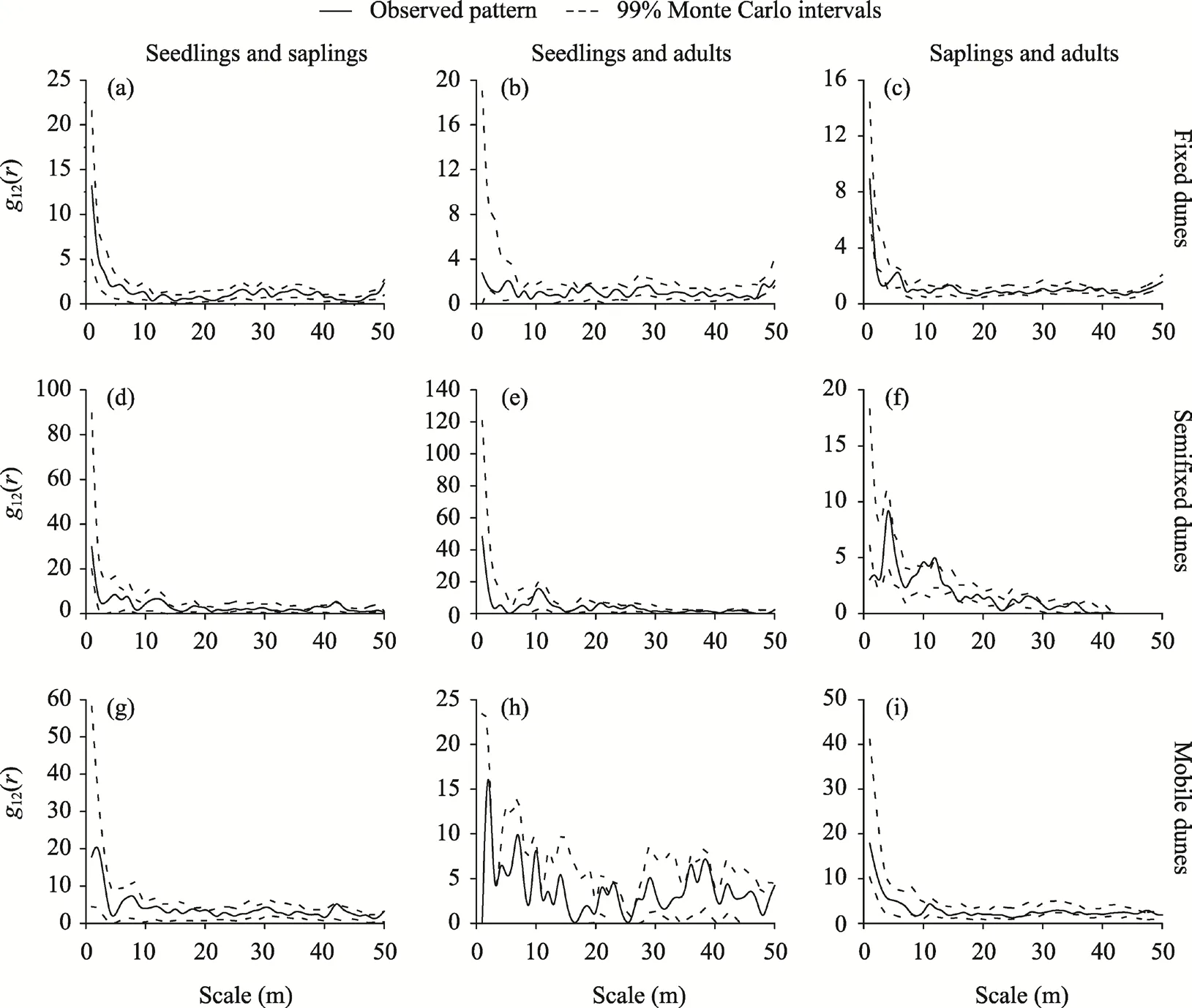
Fig. 7 Spatial association of H. persicum in different life stages in fixed dunes (a-c), semifixed dunes (d-f), and mobile dunes (g-i)
3.4 Regeneration of the seedlings and saplings of H. ammodendron and H. persicum
In the radius of 0-2 m outside the target trees, the densities of the regenerated seedlings and saplings were the largest forH. ammodendronandH. persicum, and with the increase in the radius of the target trees, the densities of the regenerated seedlings and saplings decreased (Fig.8). The average regeneration densities of seedlings and saplings ofH. ammodendronfollowed the order of fixed dunes>semifixed dunes>mobile dunes in the radius of 0-6 m outside the target trees. The density of regenerated seedlings and saplings ofH. persicumwithin a radius of 0-2 m followed the order of semifixed dunes>mobile dunes>fixed dunes, and within 2-4 m and 4-6 m, the density followed the order of mobile dunes>fixed dunes>semifixed dunes.
Correlation analysis was performed on the regenerated seedlings, sapling densities, and competition indices ofH. ammodendronandH. persicumin fixed dunes, semifixed dunes, and mobile dunes (Table 3). Within a radius of 0-2 m, the regenerated seedlings and sapling densities ofH. ammodendronandH. persicumwere significantly positively correlated with the competition index (P<0.01). In the radius range of 2-4 m, all regenerated seedlings and saplings ofH. ammodendronwere positively correlated with the competition index (P<0.05),and the density of surviving regenerated plants was not associated with the competition index.In the radius range of 2-4 m, the density of all regenerated seedlings and saplings ofH.persicumhad no relationship with the competition index, and the densities of surviving and regenerated seedlings and saplings had significant correlations with the competition index(P<0.01). In the radius range of 4-6 m, the regenerated seedlings and sapling densities ofH.ammodendronandH. persicumwere not related to the competition index.
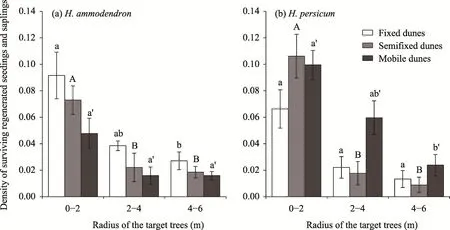
Fig. 8 Density of surviving regenerated seedlings and saplings in the different radius of target trees for H.persicum (a) and H. persicum (b) in fixed dunes, semifixed dunes, and mobile dunes. Different lowercase letters indicate significant differences in the density of surviving regenerated seedlings and saplings of the same population in fixed dunes at different circle radii (P<0.05). Different capital letters indicate significant differences in the density of surviving regenerated seedlings and saplings of the same population in semifixed dunes at different circle radii (P<0.05). Different apostrophe lowercase letters indicate significant differences in the density of surviving regenerated seedlings and saplings of the same population in mobile dunes at different circle radii (P<0.05). Error bars indicate the standard deviation.

Table 3 Correlation between the density of seedlings and saplings and the competition index for H. persicum and H. persicum
4 Discussion
4.1 Intraspecific and interspecific competition between H. ammodendron and H. persicum
The competition of plants is mainly determined by the density of tree species, individual size, and ecological habits. In this paper, the method of gradually expanding the range was used to determine that the optimal neighbor range for the competition between theH. ammodendronandH. persicumcommunities was 6 m. The reason may be that in the early stage of growth and development ofH. ammodendronandH. persicum, the niches of adjacent seedlings strongly overlap, resulting in intense intraspecific competition. With growth, the plant height and crown width increase, and the ability of individuals to obtain resources and occupy space is continuously enhanced, resulting in an increase in the optimal competition range (Zheng et al., 2017). In different habitats, the number ofH. ammodendronandH. persicumcommunities, as well as the strength of intraspecific and interspecific competition, are clearly different. Our study found that the number of plants and the intraspecific and interspecific competition intensities ofH.ammodendronfollowed the order of fixed dunes>semifixed dunes>mobile dunes, while the number of plants and the intraspecific and interspecific competition intensities ofH. persicumfollowed the order of mobile dunes>semifixed dunes>fixed dunes. Although bothH.ammodendronandH. persicumbelong to Chenopodiaceae, their suitable habitats are obviously different (Guo et al., 2005).H. ammodendronis mainly distributed in desert interdune lowlands,dry riverbeds, lake basin edges, piedmont plains, and stony gravel fields, whileH. persicummainly grows on mobile dunes or semifixed dunes (Chang et al., 2006). In this study, we discovered thatH. ammodendrongrows primarily on fixed dunes, whereasH. persicumgrows primarily on semifixed and mobile dunes. The reason may be that the soil water content of fixed dunes was higher than that of semifixed dunes and mobile dunes. The water in fixed dunes can support a larger area of vegetation, while the high soil water content can effectively alleviate the local competition relationship ofH. ammodendronand improve the survival rate ofH.ammodendronplants (Wang, 2017). In addition, the content of soil organic matter in the fixed dune increased, the nutrient content of the soil improved, and the degree of soil alkalinity was alleviated, all of which are conducive to the growth and development ofH. ammodendron.Studies have shown that the distribution characteristics ofH. ammodendronandH. persicumare closely related to their differences in water utilization (Liu, 2018). In the 0-60 cm soil surface layer,H. persicumhas a well-developed root system, and the number of lateral roots is much higher than that ofH. ammodendron.H. persicum's growth mainly depends on rainfall,regeneration, allocation, and subsurface root synergy to make the most of precipitation (Shi et al.,2016). In areas with harsh environments and away from the shade of trees,H. persicumcan achieve higher water absorption efficiency by increasing the specific surface area and specific length of fine roots (Tian and Liu, 2020). In contrast, the soil surface root system ofH.ammodendronis limited, the growth and utilization of precipitation are limited, and this species mainly tends to use groundwater (Yang et al., 2011). Therefore,H. persicumhas advantages in semifixed dunes and mobile dunes with relatively harsh natural habitat conditions, and the number of plants is relatively high, which is consistent with the findings of Wang et al. (2017) and Li et al. (2021). WhileH. ammodendronis more suitable for growing in relatively stable fixed dunes, the more stable the dunes, the higher the abundance ofH. ammodendron, the greater the intensity of intraspecific competition, and the lower the competitive pressure fromH. persicum.
Plant diameter at breast height is the main factor affecting the competition intensity (Laungani and Knops, 2009). In the same habitat, the competition intensities of theH. ammodendronandH.persicumof different size classes were different. The competition intensities of both populations followed the order of seedlings>saplings>adults, and the competition intensities gradually decreased with increasing plant diameter class. The reason may be that youngH. ammodendronandH. persicumundergo vigorous growth and development, and the demand for resources and space is greater; therefore, the competition is fierce. With increasing age, the canopy gradually sets and occupies a certain space, and the self-thinning effect also increases the distance between the plants (Huang et al., 2016). As a result, the growth ofH. ammodendronandH. persicumis relatively stable. The intensity of competition among adults in terms of light, temperature, water,and other environmental resources is gradually reduced, and the plants are subject to gradually less intraspecific and interspecific competition. Therefore, as the basal diameter increases, the intensity of competition betweenH. ammodendronandH. persicumdecreases.
In this study, we used linear, hyperbolic, power function, and logarithmic equations to perform regression fitting of the competition intensities ofH. ammodendronandH. persicum. The results showed that the correlation coefficient simulated by the power function model was the best;therefore, this equation was used as the competition intensity prediction model. The results of the prediction model showed that the competitive pressure onH. ammodendronandH. persicumdecreased as their basal diameter increased, which is consistent with the conclusions of many scholars regarding the competition of other plants (Zhang et al., 2016; Liu et al., 2020; Wang et al., 2021). When the basal diameter of the target tree exceeded 15 cm, the intrapopulation competition intensity of theH. ammodendronandH. persicumcommunities was maintained at a low level and tended to stabilize. Therefore, in the regeneration of theH. ammodendronandH.persicumnatural forests, measures such as selective cutting and artificial tending of seedlings should be implemented to strengthen the protection of the small- and medium-diameter trees.Rational management ofH. ammodendronandH. persicumnatural forests should be carried out properly before the basal diameter reaches 15 cm to reduce the competitive pressure.
4.2 Spatial pattern of H. ammodendron and H. persicum
Under natural conditions, the spatial distribution pattern of tree communities is determined by their biological characteristics, habitats, and interactions (Shen et al., 2016a). In this study, we discovered that the distribution patterns of bothH. ammodendronandH. persicumin the three different life stages showed strong random distribution and aggregated or uniform distribution patterns at some scales. However, the spatial distribution patterns ofH. ammodendronandH.persicumin Ganjia LakeHaloxylonForest Nature Reserve of China showed aggregated distribution at small scales and random or uniform distribution at larger scales (Yan et al., 2021).The reason for the difference between the two results may be the long-term adaptation and interaction between species and the environment. In this study,H. ammodendronandH. persicumgrew in the desert. The lack of water conditions restricts the survival, growth, and development of plants. The competition for water among mature plants is extremely fierce. To ensure their own growth, development, and reproduction, there was a random distribution within the population due to the competition for resources and survival space (Sun et al., 2020). In addition,H.ammodendronandH. persicumare wind-pollinated species, and their communities can reproduce and pollinate effectively (Wei et al., 2005). In this study area, the wind speed and the harsh desert environment have a strong impact on the flowering and pollination of the population. As a result,the seeds disperse farther away, and the population is extremely light-loving and does not easily form strong-scale aggregations under the parent tree (Chi et al., 2020). As the basal diameter of a plant increases, intraspecific and interspecific competition betweenH. ammodendronandH.persicumincreases, and self- and other-thinning effects occur, thus inhibiting the growth of surrounding individuals and reducing the degree of population aggregation.H. ammodendronandH. persicumshowed patterns of aggregated or uniform distribution in some scale ranges, which may be due to environmental heterogeneity. In local small environments, there may be better environmental conditions, which can support a relatively high number of small-diameter or even larger-diameterH. ammodendronandH. persicumindividuals to form an aggregated or evenly distributed pattern at a certain scale (Lv et al., 2012).
The spatial correlations of individuals at different diameter levels within a population describe the spatial distributions and functional relationships among individuals comprising a plant population at a particular time (Shields et al., 2014; Wang et al., 2016). This relationship includes not only the spatial distribution relationship but also the functional relationship between individuals, reflecting the status quo of the population and the mutual relationships within the population (Wang et al., 2010). Our results indicated that there was no correlation between the seedlings, saplings, and adults of theH. ammodendronandH. persicumcommunities, while negative and positive correlations were found in some small-scale ranges. There was no significant correlation between seedlings and saplings, adults and saplings, and adults at some scales independent of each other in the three dunes, which could be attributed to the small number of seedlings and adults and their random distribution in the sample plots and indicates that there was no significant intraspecific competition and less overlap of ecological niches at some scales.This may also be due to the large growth space ofH. ammodendronandH. persicum, both of which are strongly positive tree species. The growth of seedlings and saplings requires certain light conditions. It is difficult to obtain sufficient light under adults, as seedlings and saplings cannot survive and grow well under the canopy. They are independent of each other and reduce interdependence, which is favorable for the development of theH. ammodendron and H.persicumcommunities, and has great significance for the stable growth of the entire population(Zhang et al., 2010; Shen et al., 2016b).
There were negative correlations between seedlings and saplings, seedlings and adults, saplings and adults in theH. ammodendronat a small scale in the semifixed dunes population. The reason is that saplings and adults have strong inhibitory effects on the renewal of seedlings, and intraspecific competition is fierce. With the enhancement of saplings' ability to resist external stress, coupled with their increased demand for space resources and nutrients, competition among different age groups will continue to increase (Song et al., 2010). The difference between saplings and adults may be due to their close spatial distance and competition with each other. The positive correlation at the mesoscale indicates that the abundance of resources at the mesoscale, or subtle changes in habitat, lead to differences in the microecological niches of the species, and that seedlings, saplings, and adults can coexist harmoniously with each other (Qi, 2011). However,due to the shortage of groundwater resources, these positive relationships between saplings and seedlings and seedlings and adults shift due to competition for water resources (Feng, 2017).They end up being uncorrelated on a large scale, possibly because individuals at one growth stage of the same species create conditions for the survival of individuals at another growth stage,indicating that they have complementary requirements in the habitat (Cui, 2015).
4.3 Natural regeneration of H. ammodendron and H. persicum communities
Natural regeneration is an important ecological process in natural forest ecosystems and is the fundamental way to restore forest stands (Li et al., 2021). Good natural regeneration can optimize the structure and function of the ecosystem and form natural forests with high quality and stability(Zhang et al., 2014). In this study, we found that the regeneration densities of the seedlings and saplings of theH. ammodendronandH.persicumcommunities were the highest in the range of 0-2 m, and the regeneration densities decreased with increasing range radius. This is consistent with the results of the competition intensities ofH. ammodendronandH. persicum. With the increase in the competition index, the density of the regenerated saplings in the range of different circle radii also increased, indicating that the higher the competition index, the smaller the basal diameter, the weaker the individual's competitiveness, the lower the competition pressure of the surrounding individuals, the stronger their competitive pressure from surrounding individuals, and the less competitive pressure they can generate on surrounding regenerated seedlings and saplings, so regenerated young trees tend to be distributed around individuals with a high competition index(Pan et al., 2019). In addition, the mother trees ofH. ammodendronandH. persicumhave more individuals in the community and produce more seeds so that there are enough provenances around the mother tree (Li et al., 2009), and their seeds have a strong germination ability (Zhang et al.,2010), which results in increased regeneration density of seedlings and saplings at a smaller scale.
We found that the seedling and sapling renewal densities ofH. ammodendronin fixed dunes were higher than those in semifixed and mobile dunes, while the seedling and sapling renewal densities ofH. persicumin fixed dunes were mostly lower than those in semifixed and mobile dunes. This may be related to the stability of the dunes.H. ammodendronis more suitable for stable dunes and has a higher regeneration density. In contrast,H. persicumhas a developed root system and strong adaptability to harsh environments. It can grow and survive better in semifixed and mobile dunes environments, which leads to a high density of sapling regeneration in this environment. In addition, we found that in the radius range of 0-2 m, the densities of regenerated seedlings and saplings ofH. ammodendronandH. persicumwere significantly and positively correlated with the competition index. As the range radius increased, the densities of regenerated seedlings and saplings were not correlated with the competition index. This may be due to the greatest density of regenerated seedlings and saplings in the 0-2 m radius, the better environment in the understorey ofH. ammodendronandH. persicum, and the stronger competitiveness of the plants. With the increase in the radius range, the growth environments of seedlings and saplings changed, and the light gradually increased. These factors will have a certain impact on the germination and growth of seedlings, which is not conducive to the renewal of seedlings. The density of regenerated seedlings was obviously reduced, and the competition for water and nutrients among plants increased (Hu, 2022).
5 Conclusions
In this study, analysis methods such as the Hegyi competition index and point pattern analysis were used to reveal the characteristics of competition, spatial pattern, and regeneration ofH.ammodendronandH. persicumin different dunes and life stages.H. ammodendrondominated in the more stable fixed dunes, with significantly greater competitive intensity thanH. persicum. In contrast,H. persicumdominated in semifixed dunes and mobile dunes, where competition was more intense than that ofH. ammodendron.The competition intensity was the greatest for both the seedlings ofH. ammodendronandH. persicumin all three dunes. The distribution patterns of bothH. ammodendronandH. persicumshowed a strong random distribution in the three life stages. The seedlings, saplings, and adults ofH. ammodendronandH. persicumwere mainly uncorrelated. The regeneration density of the seedlings and saplings of theH. ammodendronandH. persicumcommunities was the highest in the range of 0-2 m. These findings may help to improve predictions of the development of plant communities in the Gurbantunggut Desert.
Acknowledgements
This study was funded by the Open Project of Xinjiang Laboratory of Lake Environment and Resources in Arid Zone (XJDX0909-2022-4) and the PhD Early Development Program of Xinjiang Normal University(XJNUBS2113).

- Journal of Arid Land的其它文章
- Antelope adaptations to counteract overheating and water deficit in arid environments
- Leaf morpho-physiology and phytochemistry of olive trees as affected by cultivar type and increasing aridity
- Leaf stoichiometry of Leontopodium lentopodioides at high altitudes on the northeastern Qinghai-Tibetan Plateau, China
- Manipulated precipitation regulated carbon and phosphorus limitations of microbial metabolisms in a temperate grassland on the Loess Plateau, China
- Effects of native and invasive Prosopis species on topsoil physiochemical properties in an arid riparian forest of Hormozgan Province, Iran
- Contents and spatial distribution patterns of heavy metals in the hinterland of the Tengger Desert, China