
Effect of mini-cutting size on adventitious rooting and morphophysiological quality of Ilex paraguariensis plantlets
2021-03-29 09:19:34NathaliaPimentelDeniseGazzanaJanainadeFatimaSpanevelloKelenHaygertLencinaDilsonAntnioBisognin
Journal of Forestry Research 2021年2期
Nathalia Pimentel · Denise Gazzana · Janaina de Fatima Spanevello · Kelen Haygert Lencina · Dilson Ant?nio Bisognin
Abstract As the energizing and therapeutic effects of infusions of mate ( Ilex paraguariensis A.St.-Hil) leaves are increasingly recognized, rising global consumption is also increasing the demand for plantlets with high genetic and morphological qualities. Mini-cuttings are an alternative for clonal plantlets; however, more details are needed on factors that affect the success of this technique. Here, toward maximizing production, we evaluated adventitious rooting competence of mini-cuttings of different sizes and the morphophysiological quality of the resultant plantlets. Small (one bud; length up to 2.5 cm), medium (multiple buds; length 2.6–5.0 cm), and large (multiple buds; length 5.1–10.0 cm) mini-cuttings were planted in a mixture of commercial substrate, vermiculite, and coarse sand (1:1:1 v:v:v), grown in a humidity chamber, then evaluated after 45, 60, 75, and 90 days for survival, percentage of callus and shoot formation, number and length of shoots, percentage of rooted cuttings, and number and length of roots. Any rooted mini-cuttings were then cultivated in commercial substrate and subsurface soil (2:1 v:v) and evaluated after 30, 60, 90 and 120 days of cultivation for morphophysiology of the plantlets, percentage of survival, number of leaves, shoot height, stem diameter, and ratio of shoot height to stem diameter. After 120 days, the plantlets were evaluated for the ratio of shoot and root dry mass, total length, surface area, total volume, and mean diameter of roots and Dickson quality index. Adventitious rooting of the mini-cuttings and the morphophysiological quality of the plantlets were not influenced by the size of the vegetative propagule. Therefore, single-bud mini-cuttings have similar adventitious rooting competence, and result in plantlets with adequate morphophysiological quality, based upon leaf development, shoot height, stem diameter and length, and surface area, volume and diameter of roots. Single-bud mini-cuttings up to 2.5 cm long can thus be used to maximize production of mate plantlets by mini-cuttings.
Keywords Aquifoliaceae · Clonal forestry · Mini-cutting technique · Plantlet production · Vegetative propagation
Introduction
Among the native tree species explored in South America, mate ( Ilex paraguariensis A.St.-Hil; Aquifoliaceae) is one of greatest in socioeconomic, environmental, and cultural importance. The leaves are used for beverages, popularly known as chimarr?o and tereré, as a source of caffeine to replace tea and coffee or as a therapeutic agent (Bracesco et al. 2011). The therapeutic effect of the leaves is due to the presence of polyphenols (Gugliucci et al. 2009), especially flavonoids, tannins, chlorogenic acid (Bastos et al. 2007; Menini et al. 2007), vitamins, and saponins (Bastos et al. 2007; Menini et al. 2007), that help improve memory (Nabi et al. 2013), promote cardioprotection (Cahuê et al. 2019), reduce weight (Antunes et al. 2017) and LDL cholesterol levels and act as an antioxidant and anti-inflammatory agent (Bracesco et al. 2011).
Evidence of the innumerable benefits of mate leaf infusions has increased the demand for raw material and interest in establishing or renewing plantations, maximizing the effi ciency of land use and, consequently, increasing rural income (Antoniazzi et al. 2018). Plantlets with well-developed aerial parts and root systems are required for homogenous plantations with high leaf productivity (Pimentel et al. 2017).
An alternative for the production of plantlets of this species is the mini-cutting technique, which uses vegetative propagules produced from either asexually propagated plantlets or seminal seedlings that are cultivated in a humidity chamber for adventitious root formation (Sá et al. 2018). Although the technique has been tested for producing mate plantlets (Wendling et al. 2007; Brondani et al. 2007, 2008; Nagaoka et al. 2013; Kratz et al. 2015; Sá et al. 2018; Pimentel et al. 2019 ), the most important factors that affect adventitious rooting of the mini-cuttings are not well known, and the technique still needs to be optimized for effi cient production.
In general, several factors, such as rejuvenation of the stock plants, the time of the year the collection is performed, application of phytoregulators, nutrition (Xavier et al. 2013), and the size of the vegetative propagules collected in the mini-clonal hedge (Engel et al. 2019) are known to influence adventitious rooting competence. The ideal size varies according to the plant species and vegetative propagation technique, inducing great variation in the rhizogenic process and in the development of vegetative propagated plantlets (Vigl and Rewald 2014). The present study was thus aimed at evaluating different sizes of mini-cuttings for adventitious rooting competence and the morphophysiological quality of the resultant plantlets to maximize production of mate plantlets from mini-cuttings.
Materials and methods
The experiments were conducted from July 2016 to March 2017 in the greenhouse at the Center for Plant Breeding and Vegetative Propagation (29° 43′ 20″ S, 53° 43′ 11″ W), Department of Plant Science of the Federal University of Santa Maria, Santa Maria, Rio Grande do Sul, Brazil. The plant material consisted of 6-month-old mini-stumps from 14 clones derived from in vitro germination of zygotic embryos (clones 06SM17, 06SM15, and 06SM12) (Pimentel et al. 2019), from cuttings ofepicormic shoots of adult trees (clones 10SM07, 13SMU05, 13SMU01, and 13SMI62) and from cuttings of seminal seedlings of mate (clones 14SMC02, 14SMC03, 15SMI04, 15SMI02, 15MA05, 15MA07, and 15MA03).
The mini-stumps were established in a mini-clonal hedge with a closed soilless system composed of a 6-mm-thick fibrocement tile suspended on iron trestles. The tile surface was coated with a transparent plastic film 100 μm thick, followed by a layer of approximately 3 cm of medium gravel. An anti-aphid screen was placed on the gravel before a layer of approximately 6 cm of coarse sand. The miniclonal hedge was fertigated daily for 15 min at a flow of 8.67 L min?1, with a nutrient solution (in mg L?1: 117.0 N in nitrate form; 15.75 N in ammonium form; 14.63 of P; 131.62 of K; 84.0 of Ca; 25.21 of Mg; 73.28 of S; 0.01 of B; 0.02 of Cu; 69.73 of Fe; 0.03 of Mn; 0.008 of Zn; 0.0016 of Mo). The pH of the nutrient solution was maintained between 5.5 and 5.8 and electrical conductivity kept at 1.5 dS m?1.
Sprouts from the mini-stumps of mate were collected in the winter (July 2016), according to the best rhizogenic responses verified by Pimentel et al. ( 2019). These sprouts were sectioned into mini-cuttings of different sizes: small (one bud; cutting length up to 2.5 cm), medium (multiple buds; length 2.6–5.0 cm) and large (multiple buds; length 5.1–10.0 cm). The small mini-cuttings were cut from portions of the sprouts without an apical bud; the medium and large cuttings were obtained from longer portions with the apical bud. After leaf area on the mini-cuttings was reduced by 50%, the cuttings were cultivated in polystyrene trays containing a mixture of pine bark-based commercial substrate, vermiculite, and coarse sand (1:1:1 v:v:v) without auxin treatment, as described by Pimentel et al. ( 2019). The chemical and physical characteristics of substrate composition were pH in H2O = 5.40; electrical conductivity = 0.46 mS cm?1; dry density = 878.84 kg m?3; total porosity = 62.02%; aeration space = 28.00%; remaining water = 26.37% and available water = 7.65%.
The trays of mini-cuttings were maintained in a humidity chamber in a greenhouse at the Center for Plant Breeding and Vegetative Propagation. The humidity chamber was overlaid with a transparent plastic film and protected with 50% nylon shading, with relative air humidity of approximately 85%, provided by automated nebulization 8 times a day for 1 min. After 45, 60, 75, and 90 days, the minicuttings were evaluated for percentage of survival and callus and shoot formation, number and length of shoots, percentage with rooting, and number and length of roots. The minicuttings were considered rooted when they had at least one adventitious root that was ≥ 0.1 cm long. The experimental design was completely random, with 10 repetitions of 7–15 mini-cuttings each, based on the productivity of sprouts on the mini-stumps.
After 90 days in the humidity chamber, the rooted minicuttings ofeach of treatment were grown in 500-cm 3 polyethylene bags (10 cm wide × 13 cm long) (Pimentel et al. 2017) containing a mixture of a commercial pine-barkbased substrate and subsurface soil (2:1 v:v) to evaluate the morphophysiological quality of the plantlets. The chemical and physical characteristics of the substrate were pH H2O = 4.97; electrical conductivity = 0.89 mS cm?1; dry density = 590.23 kg m?3; total porosity = 73.51%; aeration space = 20.79%; remaining water = 33.73% and available water = 20.27%. The plantlets were arranged on benches with 70% of total incident radiation (approximately 7805 lx) because they were protected with 30% nylon shading throughout the experiment. Plants were irrigated daily with a hand irrigator (3.6 L m?2). The experiment was conducted in a completely random design, with five repetitions of 4–20 plantlets each, according to the number of mini-cuttings rooted in the previous experiment.
The mate plantlets were evaluated after 30, 60, 90, and 120 days for survival percentage, number of leaves, shoot height (cm), stem diameter (mm), and the ratio of shoot height to stem diameter. Shoot height was measured from the substrate to the apex of the last-formed leaf, and stem diameter was measured at the substrate level with a digital caliper.
After 120 days, four random plantlets for each treatment were collected to determine dry mass of the shoot and roots and analyze the morphophysiology of the root system. The plantlets were washed in tap water and cut at the substrate line to separate shoots and roots, which were then placed in separate paper bags dried in an oven at 105 °C until constant mass. Roots were then scanned in an Epson 11000XL scanner and analyzed using WinRhizo Pro software (Regent Instruments, Canada) to determine total length, surface area, and volume and average diameter of the roots. In addition, we estimated the ratio of shoot dry mass to root dry mass and calculated the Dickson quality index (DQI) using the following equation (Dickson et al. 1960):

where TDM is the total dry mass of the plantlets (g), SH is the shoot height (cm), SD is the stem diameter (mm), and RSR is the ratio of shoot dry mass to root dry mass.
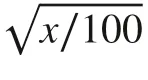
Results and discussion
In this study, the size of the mate mini-cuttings had a significant effect on the percentage of survival and callus formation after 60 days ( p = 0.0025 and 0.0125, respectively), 75 ( p = 0.0000 and 0.0002, respectively) and 90 days ( p = 0.0048 and 0.0067, respectively) in the humidity chamber (Fig.1 a, b). The high survival of the medium-sized and the large mini-cuttings (Fig.1 a) suggests that the air humidity provided by nebulization was adequate for the survival ofeven the largest vegetative propagules, which obviously require more water (Lima et al. 2006). For the vegetative propagation of the tree clones Populus ‘Max 4′ and Salix ‘Inger’, the longest cuttings had the highest survival during rhizogenesis (Vigl and Rewald 2014), as also found in the present study with mate mini-cuttings. For the percentage of callus formation (Fig.1 b), the superiority of medium and large mini-cuttings may have been due to the higher number of buds and leaves on the vegetative propagules than on the small mini-cuttings. The leaves synthesize soluble sugars, which provide energy for callogenesis (Shao et al. 2018). Greater callogenesis was also found for vegetative propagules of Varronia curassavica Jacq. that had more leaves (Bischoff et al. 2017).
The size of the mate mini-cuttings also had a statistically significant effect on the percentage of shoot formation after 45, 60, 75, and 90 days of cultivation ( p = 0.0035, 0.0001, 0.0144, 0.0415, respectively) (Fig.1 c), but did not influence the number and length of shoots ( p = 0.1015 and 0.4292 [45 days], 0.3326 and 0.4178 [60 days], 0.1391 and 0.0994 [75 days], 0.4885 and 0.3111 [90 days]) (Fig.1 d, e). The shoot formation was greatest on the small mini-cuttings (Fig.1 c), which lacked the apical bud and were thus not subject to apical dominance, perhaps increasing the concentration of cytokinins in the lateral buds (Taiz and Zeiger 2013) and thus promoting the growth of lateral shoots.
The ideal size for vegetative mini-cuttings with regard to adventitious rooting varies with the forest species. Short cuttings had higher competence for adventitious rooting of Melaleuca alternifolia Cheel (Oliveira et al. 2008), but larger vegetative propagules were best for Picea abies (L.) Karst. (Ouyang et al. 2015) and Handroanthus heptaphyllus (Vell.) Mattos (Rodrigues et al. 2017). For mate, cuttings from adult plants that are longer than 10 cm are more proficient for adventitious rooting (Duarte et al. 2019), butthe ideal size of mini-cuttings for mate has not been known; vegetative propagules between 2 and 7 cm have been used to study adventitious rooting (Wendling et al. 2007; Brondani et al. 2007, 2008; Nagaoka et al. 2013; Kratz et al. 2015; Sá et al. 2018; Pimentel et al. 2019).
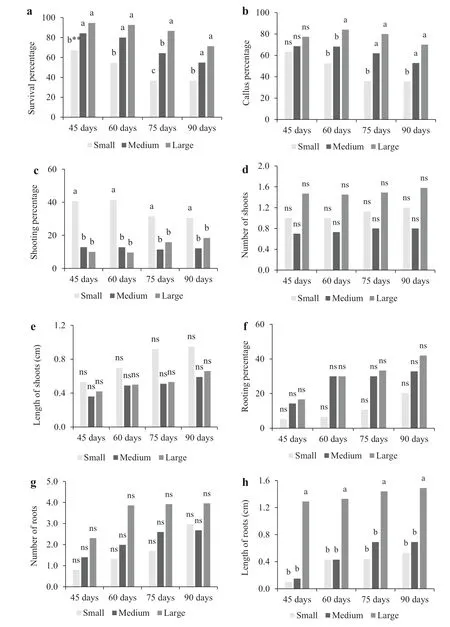
Fig.1 Percentage of survival and callus and shoot formation, number and length of shoots, percentage of rooted minicuttings, and number and length of roots on mate mini-cuttings of different sizes after 45, 60, 75, and 90 days of cultivation. Sizes: small mini-cuttings (one bud, mini-cutting up to 2.5 cm long), medium (multiple buds, 2.6–5.0 cm long) and large (multiple buds, 5.1 to 10.0 cm long). **Lower case letters indicate means differed significantly among mini-cutting size for a particular time ofevaluation according to Scott–Knott’s test at the 5% significance level. ns Not significant according to the F-values in ANOVA
In the present study, the different sizes of mate minicuttings did not differ significantly in relation to the rooting percentage and number of roots after 45, 60, 75, and 90 days in the humidity chamber ( p = 0.4649 and 0.4202, 0.1212 and 0.1624, 0.0686 and 0.1884, 0.2432 and 0.3035, respectively; Fig.1 f, 1g; Fig.2 a–c). This result was probably due to the larger mini-cuttings having more buds than on the small (one bud). The bud can act as a heavy sink and consumer of reserves (Lima et al. 2006), resulting in vegetative propagules of different sizes but with similar amounts of photoassimilates. The photoassimilates are synergistic substances for adventitious rooting and may allow similar reserves to achieve similar rhizogenic rates on the three sizes of mate mini-cuttings. A similar lack of influence of vegetative propagule size on rhizogenesis was found for Allanblackiafloribunda Oliv. (Atangana and Khasa 2008) and Santalum austrocaledonicum Vieill. (Tate and Page 2018).

Fig.2 Rooted mini-cuttings of mate a– c after 90 days in a humidity chamber and plantlets d– f after 120 days in the greenhouse. Mini-cuttings of small ( a, d), medium ( b, e), and large ( c, f) sizes are shown. Bar = 2 cm
Adventitious roots were longest on large mini-cuttings after 90 days of cultivation in the humidity chamber (Figs. 1 h, 2 c). Duarte et al. ( 2019) also verified that 10 cm cuttings of adult mate plants had longer roots after 120 days; however, they only used nonrejuvenated propagules. Their result suggests superior survival of plantlets from larger vegetative propagules, which was not observed in the present study (Fig.3). The survival percentage after 60, 90, or 120 days in the greenhouse was not influenced ( p = 0.4221, 0.6634, 0.6085, respectively) by the size of the original mini-cutting (Fig.3 a). According to Fachinello et al. ( 2005), the number of roots formed on vegetative propagules (Fig.1 g) has greater influence than the length of the roots (Fig.1 h) on survival of the new plantlets. Thus, the similar survival rate of mate plantlets (Fig.3 a) may be associated with the similar number of roots formed during the rhizogenic process in the mini-cuttings of different sizes (Fig.1 g).
Plantlets from large mini-cuttings had more leaves (Fig.3 b) and longer shoots (Fig.3 c) compared with the other mini-cutting sizes after 30 days in the greenhouse. However, by 120 days, the mate plantlets from mini-cutting did not differ significantly ( p = 0.1567 and 0.0981, respectively) for these traits (Fig.2 d–f; Fig.3 b, c). In general, the longer the vegetative propagule, the greater the carbohydrate reserves (Fachinello et al. 2005), allowing more vigorous initial growth of plantlets from this size of propagule. However, these differences were not found by the end of the evaluation period.
The different sizes of mini-cutting plantlets yielded significantly different stem diameters after 30, 60, 90, and 120 days in a greenhouse ( p = 0.0001, 0.0050, 0.0085, 0.0046, respectively). The medium mini-cuttings yielded a smaller stem diameter for plantlets than did the smaller and larger mini-cuttings at 30, 60 and 90 days of cultivation (Fig.3 d). However, by the end of the experiment (120 days), the small mini-cuttings had produced plantlets with the greatest stem diameter (average 2.80 mm; Fig.3 d). This result again is likely due to the lack of apical bud on the small mini-cuttings, increasing the availability of nutrients and cytokinins to the lateral buds, and thus stimulating the growth in diameter (Ono et al. 2004). The removal of the apical bud also allowed a greater diameter increase in plants of Carica papaya L. (Ono et al. 2004). Stem diameter is considered the best morphological predictor of future plantlet development, having the highest correlation with survival rate and plant growth in the field (Haase 2008). The ideal range for stem diameter, however, varies by tree species, and no information is yet available for optimal values for mate plantlets. For the genus Eucalyptus, a minimum diameter of 2.0 mm is recommended for plantlets before transplanting in the field (Xavier et al. 2013). Thus, considering the ideal range used for Eucalyptus tree species, only mate plantlets produced from small mini-cuttings would be considered suitable for planting (Fig.3 d).
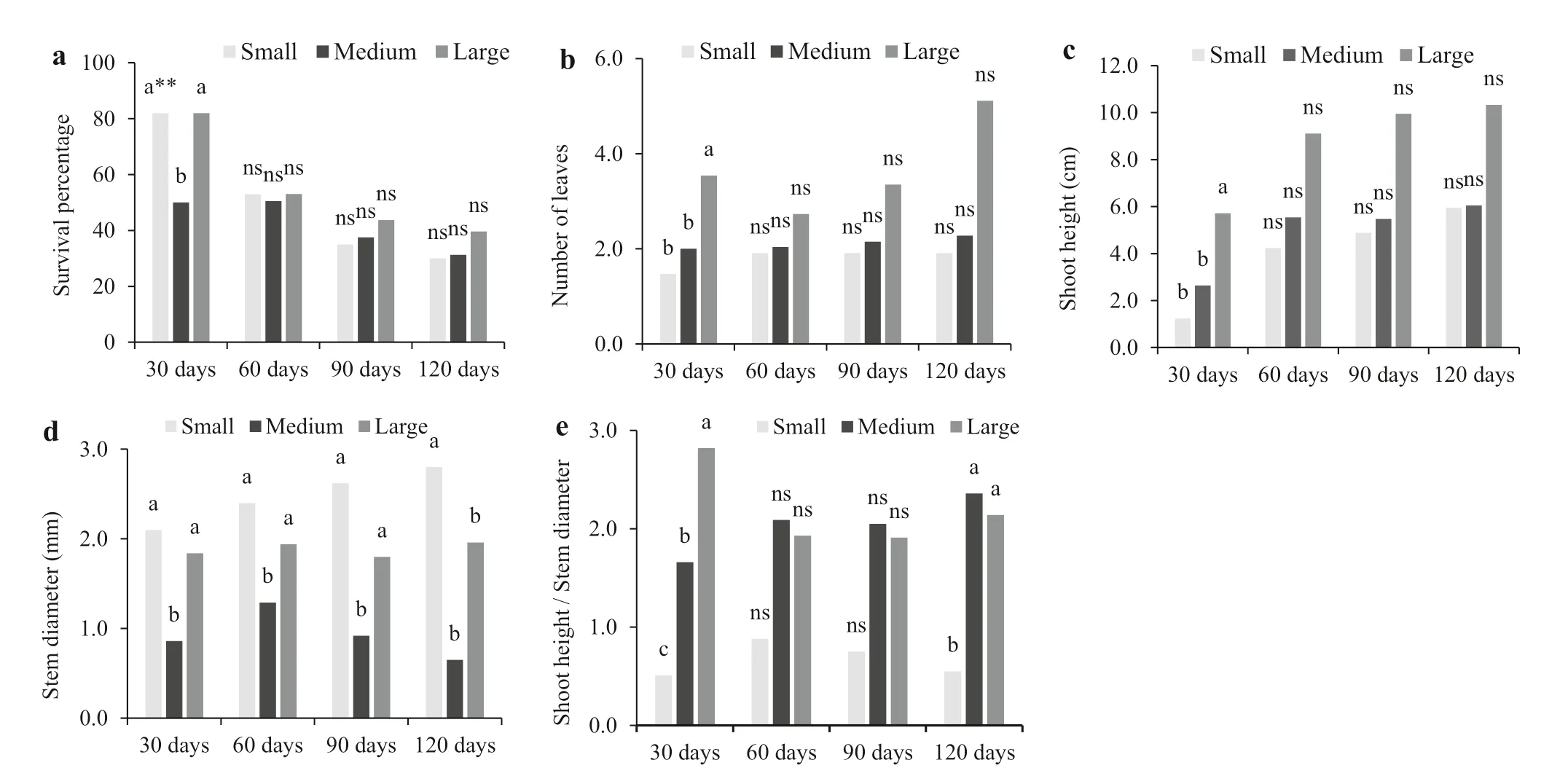
Fig.3 Percentage survival, number of leaves, shoot height, stem diameter, and shoot height to stem diameter ratio of mate plantlets from mini-cuttings of different sizes after 30, 60, 90, and 120 days in a greenhouse. *Small mini-cuttings (one bud; up to 2.5 cm long), medium (multiple buds; 2.6–5.0 cm), and large (multiple buds; 5.1–10.0 cm). **Lower case letters indicate that means differed significantly among mini-cutting size for a particular time ofevaluation in Scott–Knott’s test at the 5% significance level. ns Not significant according to the F-values in ANOVA
The use of medium-sized and large mini-cuttings led to a higher shoot height to stem diameter ratio in mate plantlets after 120 days in the greenhouse (Fig.3 e). However, regardless of the size of the initial mini-cuttings, all plantlets had a shoot height to stem diameter ratio less than 10, which according to Birchler et al. ( 1998), indicates balanced shoot development and stem diameter of plants produced from mini-cuttings.
The final plantlets generated by the different sizes of mini-cutting after 120 days in the greenhouse did not differ in the ratio of shoot to root dry mass ( p = 0.0640) (Fig.S1 a), total length, surface area, total volume, and average diameter of the roots ( p = 0.2741; 0.2666; 0.2398; 0.1154, respectively; Fig.S1 b–e). Thus, the results show a trend for root growth similar to that observed in adventitious root formation. Therefore, this trend can also be explained by a reduction in photoassimilates in the longer mini-cuttings due to their multiple buds, which probably equaled the reserve content of small mini-cuttings and, consequently, enabled satisfactory development of the root system. This result may indicate that these plantlets will grow well in the field, since a positive relationship between the quality of the root system and growth after planting was reported in 78% of the studies analyzed by Grossnickle and MacDonald ( 2018).
The Dickson quality index (DQI) is considered to be a good indicator of plantlet quality, since it uses both the robustness (ratio of shoot height to stem diameter) and the plant biomass balance (ratio of shoot to root dry mass) (Fonseca et al. 2002). Mate plantlets from medium and large mini-cuttings had the highest DQI values (0.19 and 0.12 respectively), differing statistically from those from small vegetative propagules (0.09) after 120 days (Fig.S1 f). For Pseudotsuga menziesii (Mirbel) Franco and Picea abies (L.) Karst., DQI equal to or greater than 0.20 was recommended for the quality of plantlets to be considered satisfactory (Hunt 1990). However, in the present study, the DQI values for plantlets from all sizes of mini-cuttings were lower than 0.2, which led us to consider that for mate, this value is inappropriate. DQI values were also lower than 0.2 for seedlings of native species such as Copaifera langsdorffi i Desf. (Reis et al. 2016) and Chamaecrista desvauxii var. latistipula (Delarmelina et al. 2015). These results corroborate the findings of Leles et al. ( 2006), who stated that species of the Brazilian flora need to be studied to more precisely determine index optimal for Brazilian species.
The results of the present study indicate that neither the rhizogenesis in the mini-cuttings nor the morphophysiological quality of the mate plantlets were influenced by the size of the initial vegetative propagule. Thus, the use of small mini-cuttings (with one bud and length up to 2.5 cm) is suggested for producing mate plantlets with satisfactory adventitious rooting and shooting rates in the humidity chamber and, consequently, to obtain plantlets with adequate morphophysiological quality, as evidenced by leaf number, shoot height, stem diameter and total length, surface area, total volume, and average root diameter. In addition, the use of mini-cuttings with one bud maximizes production, since numerous one-bud cuttings can be cut from one sprout. Thus, from each mini-stump in the mini-clonal hedge, more plantlets can be produced, reducing the area required for a mini-clonal hedge. Also, by using a small vegetative propagule with one bud, the time interval between collections can be increased, so that the collection can be planned according to the availability of space in the humidity chamber, maximizing the use ofone of the most costly plantlet production environments. This ability to manage the interval between collections also allows the process to be adjusted when demand is greater for mate plantlets of adequate morphophysiological quality.
Conclusions
The size of mate mini-cuttings does not affect adventitious rooting or the morphophysiological quality of the plantlets. The use of single-bud mini-cuttings up to 2.5 cm long maximizes production of mate plantlets by mini-cuttings and minimizes costs.
References
Antoniazzi MS, Rocha LD, Souza EL, Guerra D, Silva DM, Redin M (2018) Análise da cultura da erva-mate como alternativa social, econ?mica e ambiental para comunidades rurais. Ver Extem Foco 15:108–119
Antunes A, Cardoso DS, Paula CS (2017) Efeito da Ilex paraguariensis A. St. Hil. (erva mate) no controle da obesidade. Vis?o Acadêmica 18(1):14–26.
Atangana AR, Khasa DP (2008) Preliminary survey of clonal variation in rooting of Allanblackia floribunda leafy stem cuttings. Can J For Res 38:10–15
Bastos DHM, Oliveira DM, Matsumoto RLT, Carvalho PO, Ribeiro ML (2007) Yerba mate: pharmacological properties, research and biotechnology. Med Aromat Plant Sci Biotechnol 1:37–46
Birchler T, Royo A, Pardos M (1998) La planta ideal: revision del concepto, parametros definitorios e implementaction practica. Investigacion Agraria: Sistemas y Recursos Forestales 7(1/2):109–121
Bischoff AM, Vendramim DW, Gomes EM, Ribas KCZ, Engel ML, Maggioni RA (2017) Enraizamento de estacas de erva-baleeira em fun??o de diferentes concentra??es de ácido indolbutírico e número de folhas. Cienc Agrovet 16(1):41–47
Bracesco N, Sanchez AG, Contreras V, Menini T, Gugliucci A (2011) Recent advances on Ilex paraguariensis research: Minireview. J Ethnopharmacol 136(3):378–384
Brondani GE, Wendling I, Santin D, Benedetti EL, RovedaLF OA (2007) Ambiente de enraizamento e substratos na miniestaquia de erva mate. Sci Agrar 8(3):257–267
Brondani GE, Araujo MA, Wendling I, Kratz D (2008) Enraizamento de miniestacas de erva-mate sob diferentes ambientes. Pesqui Florest Bras 57:29–38
Cahuê F, Nascimento JHM, Barcellos L, Salerno VP (2019) Ilex paraguariensis, exercise and cardioprotection: a retrospective analysis. J Funct Foods 53:105–108
DelarmelinaWM CMVW, Faria JCT, Lacerda LC (2015) Uso de resíduo organico em substrato para produ??o de Chamaecrista desvauxii (Collad.) Killip var. latistipula (Benth.). Cerne 21(3):429–437
Dickson A, Leaf AL, Hosner JF (1960) Quality appraisal of white spruce and white pine seedling stock in nurseries. Forest Chron 36:10–13
Duarte MM, Mireski MC, Oliszeski A, Wendling I, Stuepp CA (2019) Rooting of yerba mate cuttings with different lengths. Rev Elet Cient da UERGS 5(1):5–11
Engel ML, Ikeda A, Fl?res PC Jr, Alcantara GB, Higa AR (2019) Influence of stem cutting position, leaves condition and size of minicuttings in rooting of black wattle. Agric For 65:191–199
Equipe Estatcamp (2014) Software action. Estatcamp- Consultoria em estatística e qualidade, S?o Carlos-SP, Brasil. https://www.porta lacti on.com.br/ .Available 02.05.19
Fachinello JC, Hoff mann A, Nachtigal JC (2005) Propaga??o de plantas frutíferas. Embrapa Informa??o Tecnológica, Brasília
Ferreira DF (2011) Sisvar: a computer statistical analysis system. Cienc Agrotecnol 35(6):1039–1042
Fonseca EP, Valéri SV, Miglioranza E, Fonseca NAN, Couto L (2002) Padr?o de qualidade de mudas de Trema micrantha (L.) Blume produzidas sob diferentes períodos de sombreamento. árvore 26(4):515–523
Grossnickle SC, MacDonald JE (2018) Why seedlings grow: influence of plant attributes. New For 49:1–34
Gugliucci A, Bastos DH, Schulze J, Souza MF (2009) Caffeic and chlorogenic acids in Ilex paraguariensis extracts are the main inhibitors of AGE generation by methylglyoxal in model proteins. Fitoterapia 80(6):339–344
Haase DL (2008) Understanding forest seedling quality: measurements and interpretation. Tree Plant 52(2):24–30
Hunt GA (1990) Effect of styroblock design and Cooper treatment on morphology of conifer seedlings. United States Departament of Agriculture, Forest Service, Fort Collins
Kratz D, Pires PP, Stuepp CA, Wendling I (2015) Produ??o de mudas de erva-mate por miniestaquia em substratos renováveis. Floresta 45(3):609–616
Leles PSS, Lisboa AC, Neto SNO, Grugiki MA, Ferreira MA (2006) Qualidade de mudas de quatro espécies florestais produzidas em diferentes tubetes. Floresta e Ambient 13(1):69–78
Lima RLS, Siqueira DL, Weber OB, Cazetta JO (2006) Comprimento de estacas e parte do ramo na forma??o de mudas de aceroleira. Rev Bras Frutic 28(1):83–86
Menini T, Heck C, Schulze J, Mejia E, Gugliucci A (2007) Protective action of Ilex paraguariensis extract against free radical inactivation of paraoxonase-1 in high-density lipoprotein. Planta Med 73:1141–1147
Nabi NU, Neeraj K, Ravi K, Preeti K (2013) Natural remedies for improving learning and memory-review. Int J Pharm 3(2):161–165
Nagaoka RE, Peres FSB, Carmo ALM, Garcia FAO (2013) Efeito do AIB no desenvolvimento de mudas clonais em genótipos de ervamate. Enciclopédia Biosfera 9(17):1182–1191
Oliveira Y, Silva ALL, Pinto F, Quoirin M, Biasi LA (2008) Comprimento das estacas no enraizamento de melaleuca. Sci Agrar 9(3):415–418
Ono EO, Grana Júnior JF, Rodrigues JD (2004) Reguladores vegetais na quebra da dominancia apical de mamoeiro ( Carica papaya L.). Rev Bras Frutic 26(2):348–350
Ouyang F, Wang J, Li Y (2015) Effects of cutting size and exogenous hormone treatment on rooting of shoot cuttings in Norway spruce [ Picea abies (L.) Karst.]. New For 46:91–105
Pimentel N, Lencina KH, Pedroso MF, Somavilla TM, Bisognin DA (2017) Morphophysiological quality of yerba mate plantlets produced by mini-cuttings. Semin Cienc Agrar 38(6):3515–3528
Pimentel N, Lencina KH, Kielse P, Rodrigues MB, Somavilla TM, Bisognin DA (2019) Produtividade de minicepas e enraizamento de miniestacas de clones de erva-mate ( Ilex paraguariensis A. St.-Hil.). Ciencia Florestal 29(2):559–570
Reis SM, Marimon-Junior BH, Morandi OS, Oliveira-Santos C, Oliveira B, Marimon BS (2016) Desenvolvimento inicial e qualidade de mudas de Copaifera langsdorffi i Desf. sob diferentes níveis de sombreamento. Ciencia Florestal 26(1):11–20
Rodrigues MB, Pimentel N, Lencina KH, Kielse P, Bisognin DA (2017) Enraizamento de miniestacas de ipê-roxo ( Handroanthus heptaphyllus Vell. Mattos). Rev Bras Pl Med 19(1):129–137
Sá FP, Portes DC, Wendling I, Zuffellato-Ribas KC (2018) Miniestaquia de erva-mate em quatro épocas do ano. Ciencia Florestal 28(4):1431–1442
Shao F, Wang S, Huang W, Liu Z (2018) Effects of IBA on the rooting of branch cuttings of Chinese jujube ( Zizyphus jujuba Mill.) and changes to nutrients and endogenous hormones. J For Res 29(6):1557–1567
Taiz L, Zeiger E (2013) Plant Physiology. Sinauer Associates, Sunderland
Tate HT, Page T (2018) Cutting propagation of Santalum austrocaledonicum: the effect of genotype, cutting source, cutting size, propagation medium, IBA and irradiance. New For 49:551–570
Vigl F, Rewald B (2014) Size matters? The diverging influence of cutting length on growth and allometry of two Salicaceae clones. Biomass Bioenerg 60:130–136
Wendling I, Dutra LF, Grossi F (2007) Produ??o e sobrevivência de miniestacas e minicepas de erva-mate cultivadas em sistema semihidrop?nico. Pesq Agropec Bras 42(2):289–292
Xavier A, Wendling I, Silva RL (2013) Silvicultura clonal: princípios e técnicas. Editora UFV, Vi?osa
 Journal of Forestry Research2021年2期
Journal of Forestry Research2021年2期
- Journal of Forestry Research的其它文章
- Stem taper functions for Betula platyphylla in the Daxing’an Mountains, northeast China
- Leaf phenotypic variation of endangered plant Tetracentron sinense Oliv. and influence of geographical and climatic factors
- Ecological variations of woody species along an altitudinal gradient in the Qinling Mountains of Central China: area-based versus mass-based expression of leaf traits
- Variations in stem radii of Larix principis-rupprechtii to environmental factors at two slope locations in the Liupan Mountains, northwest China
- A review of ecological mechanisms for management practices of protective forests
- Variation of basic density, calorific value and volumetric shrinkage within tree height and tree age of Ugandan grown Eucalyptus grandis wood