
Seed germination and seedling emergence of four tree species of southern China in response to acid rain
2021-03-29 09:19:34MatoorMohsinGilaniMulualemTigabuBoLiuTaimoorHassanFarooqMuhammadHaroonRashidMuhammadRamzanXiangqingMa
Journal of Forestry Research 2021年2期
Matoor Mohsin Gilani · Mulualem Tigabu · Bo Liu · Taimoor Hassan Farooq · Muhammad Haroon U. Rashid · Muhammad Ramzan · Xiangqing Ma
Abstract Acid rain has become a major concern due to increasing atmospheric pollution, particularly in China. We investigated whether acid rain inhibits the germination of seeds and subsequent emergence of seedling of four tree species from southern China: Cunninghamia lanceolata, Fokienia hodginisi, Pinus massoniana and Phoebe zhennan by simulating acid rain with pH of 2.5, 3.5, 4.5 and 5.5. We hypothesized that the inhibitory effect of acid rain on germination of seeds and emergence of seedling varies between species and the degree of acidity. A solution of 1 N H 2 SO 4 and 1 N HNO 3 in the ratio of 10–1 was prepared and diluted to four pH levels, and seeds were supplied with solutions of these pH values and distilled water as control and tested for germination in a controlled growth chamber. The results revealed that simulated acid rain of pH 2.5 adversely affected the germination capacity of F. hodginisi and P. zhennan; while all acid solutions significantly increased germination of P. Massoniana; but had no effect on germination of C. Lanceolata seeds. Strong acid solution (pH of 2.5) adversely affected elongation of radicle and hypocotyl as well as fresh and dry weights of radicle and hypocotyl of tender seedlings. The result demonstrated that seedling emergence is more sensitive than seed germination to simulated acid rain, and germination of conifer species are less sensitive than broad leaved species to simulated acid rain. As a whole, acid rain of pH of 3.5 is the threshold level and acid rain below this value will have a detrimental effect on seed germination and seedling emergence.
Keywords Cunninghamia lanceolata · Fokienia hodginisi · Pinus massoniana · Phoebe zhennan · Seed viability · Simulated acid rain
Introduction
Acid rain is simply defined as precipitation with a pH lower than 5.6 (Niu et al. 2010) and formed when pollutants, such as sulphur and nitrogen oxides, react with atmospheric moisture (Anitha and Ramanujan 1992). The rapid development of the Chinese economy has resulted in increased emission of atmospheric pollutants due to an increased consumption of fossil fuels. Thus, acid rain has become a serious environmental concern in China (Huang et al. 2012). Over the past three decades, the area that experienced acid rain in China had surpassed 30% of China’s land area (Xie et al. 2008; Zhang et al. 2010), which is closely related with increased fossil fuel consumption, with sulfuric acid being the major cause of acid rain (Chen et al. 2006; Dong et al. 2015). In recent years, both the acidity of the rain and the area affected by acid rain has increased dramatically in China (Huang et al. 2012).
The distribution of acid rain has shown an increasing trend in the northern, eastern and southern parts of China; and the most acid rain affected region is located south of the Yangtze River (Cheng et al. 2011). For instance, a pH of average monthly precipitation for the period 2006–2008 was below 4.5 in the Lin’an region of Zhejiang Province (Niu et al. 2010); Mei et al. ( 2010) observed an increase in the intensity of acid rain in Shanghai since 2002; and Huang et al. ( 2008) found most rainwater samples from Shanghai being acidic (pH of 2.95). Analysis of chemical characteristics of precipitation showed a strong relationship between air pollution and acid precipitation in many cities in China (Cao et al. 2009; Cheng et al. 2011; Shi et al. 2014; Wang and Han 2011).
Acid rain can cause reduction in growth and productivity of plants and other ecosystems (Britton et al. 1996; Macaulay and Enahoro 2015) through increasing soil acidity and direct foliar damage (Allen 1994; Larssen et al. 2000; Bian and Yu 1992). Among other things, acid rain could lead to foliar necrosis, abnormal metabolic processes, inhibition of seed germination and retarding seedling growth and predisposition plants to other biotic and abiotic stresses (Byres et al. 1992; Chao 1995; Fan and Wang 2000). It also exacerbates ecological risks in the agricultural ecosystem (Cui et al. 2014) through increased solubility and bioavailability of soil contaminant, such as lead (Flegal et al. 2013). In addition, acid rain poses serious human health risks, leads to corrosion of cultural relics and destroys ecosystem structures and functions; thus, becoming the major environmental problem attracting much research attention (Wang and Xu 2009; Zhang et al. 2010).
Germination of seeds and subsequent emergence of seedlings are key stages in plant population reproduction and survival, and determine the time when plants recruit into populations and communities (Rajjou et al. 2012). In order to improve current cultivation practices of tree species in southern China, we tested the effects of acid rain on germination of seeds and subsequent emergence of seedlings of four tree species: Cunninghamia lanceolata, Fokienia hodginisi, Pinus massoniana and Phoebe zhennan by simulating acid rain with different pH levels. As seeds are the primary regeneration material for large scale plantation establishment, we specifically aimed to understand how acid rain may affect early life stage processes such as seed germination and seedling growth. The species are commonly grown in southern China for a variety of purposes, ranging from timber production to soil and water conservation. The objective of the study was to determine the level of acid rain that is detrimental for establishment of the species. We hypothesized that the adverse effect of acid rain on germination of seeds and subsequent emergence of seedlings varies between species and the degree of acidity. To test this hypothesis, a factorial experiment with four species and four pH levels of simulated acid rain (2.5, 3.5, 4.5 and 5.5) and distilled water as control was conducted. Germination capacity, an estimate of the viability of a population of seeds, mean germination time as a measure of germination rate, radicle and hypocotyl length, fresh and dry weight of radicle and hypocotyl as indicators of seedling emergence were analyzed.
Materials and methods
Species description and seed samples
In this study, four commonly growing tree species in south China were used. The species used were Cunninghamia lanceolata, Fokienia hodginisi, Pinus massoniana and Phoebe zhennan. C. lanceolata (commonly known as Chinese fir) is widely planted in south China for high quality timber production. Chinese fir is also planted in parks and large gardens, and as buffer zone plantings around parking lots or along the highway. F. hodginisi (locally known as Fujian cypress), whose wild stocks are dwindling rapidly, produces high quality timber for construction purpose. Owing to its light, aromatic and resinous timber, local people highly prize the timber of this species. P. massoniana (commonly known as Mason’s pine) is a species commonly grown in productive forest plantation in southern China. The wood is used for making pulp and paper and for production of Chinese rosin (obtained from the turpentine of this pine). Leaves are often used for making special smoke flavor to a local black tea (Lapsang souchong) in Fujian Province. P. zhennan (commonly known as Gold Phoebe) is a valuable source of wood and bioactive compounds used in pharmaceutics, but it is one of the most threatened tree species in south China, included in the red list species database in China.
Seeds of all species were collected from Zhang ping Forest Farm in Fujian Province. Fujian Province has a subtropical climate with mild winter. The average temperature of the coastal regions is 7–10 °C while the hilly areas experience 6–8 °C in January. Temperatures are high during the summer, and the province often experiences typhoons coming in from the Pacific storms. The average annual precipitation is 1400–2000 mm. The seeds were processed manually and stored at 4 °C for few weeks until used for the experiment.
Simulation of acid rain
Acid solution of 10–1 ratio by chemical equivalent was prepared by mixing 1 N H2SO4 and 1 N HNO3. This ratio was in accordance with the general anion composition of rainfall in South China (Fan 1993). The stock solution was then diluted to pH values 2.5, 3.5, 4.5 and 5.5 by adding distilled water. As a control, distilled water with pH around 6.5 was used.
Germination test
The germination test was performed in the laboratory at Fujian Agriculture and Forestry University. Before sowing, seeds were disinfected by concentrated potassium permanganate solution for 20 min and then rinsed in running tap water for 5 min. Thereafter, seeds were sown in Petri dishes on filter papers (9 cm in diameter), which received 2 mL of acid solutions or distilled water. As the filter papers dried out, we added the same amount of acid solution or distilled water every other day. For each treatment, 50 seeds replicated six times were used. Germination test was run in a growth chamber set at a constant temperature of 25 ± 1 °C day and 12/12 h day/night light of 20 μE for 23 days. Germinants were counted every day when radicle protrusion reached 2 mm long. At the end of the germination test, radicle and hypocotyl length, fresh and dry weight of radicle and hypocotyl were determined. For dry weight determination, radicle and hypocotyl were oven-dried at 75 °C until constant mass.
Data analysis
The following germination parameters were calculated for each replicate in each treatment:

where GC is germination capacity, expressed in percentage, niis the number of germinated seeds at each day, N is the total number of seeds sown, MGT is the mean germination time, expressed in days, and tiis the number of days starting from the date of sowing (Bewley et al. 2013). As germination has a binomial distribution, the germination capacity data set was arcsine transformed prior to data analyses to meet the normality assumption for analysis of variance (Zar 1996). GC and MGT data sets were subjected to General Linear Model (GLM)—Univariate Analysis to determine significant differences among species and pH level of the acid solution. Tukey’s Honestly Significant Test ( P < 0.05) was used to compare significant mean differences among pH levels for each species separately. SPSS 20.0 software (SPSS Inc., Chicago, IL, USA) was used for data analysis and Origin Pro 8.5 was used for drawing all graphs.
Results
Seed germination
Significant differences in germination capacity were detected among simulated acid rain treatments, species and their interaction (Table 1). Simulated acid rain treatment of pH 2.5 significantly reduced the germination capacity compared to other treatments and the control while seeds of F. hodginisi had lower germination capacity than the other species. At individual species level, simulated acid rain had no adverse effect on germination of C. lanceolata seeds (Fig.1 a), while acid rain treatments with pH greater than 2.5 stimulated the germination of P. massoniana seeds (Fig.1 b).The germination of F. hodginisi was significantly reduced by acid rain of pH 2.5 compared to other treatments and the control (Fig.1 c). Nearly all acid rain treatments significantly reduced the germination of P. zhennan seeds (Fig.1 d).
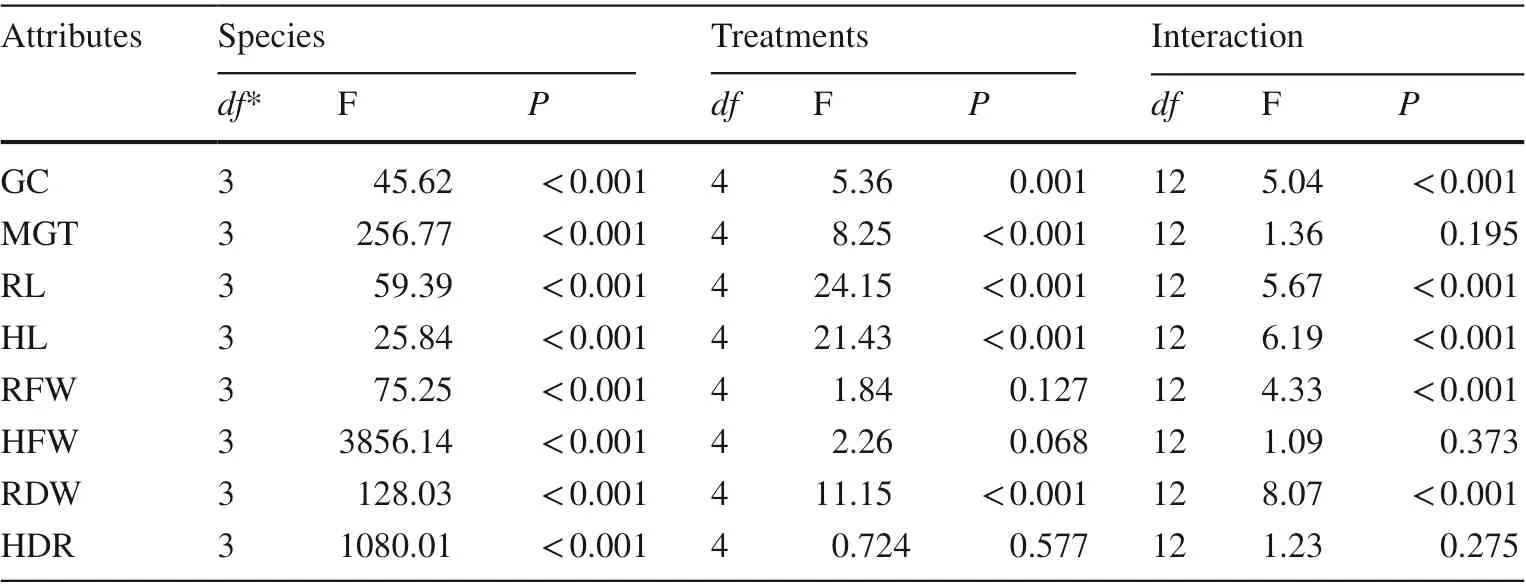
Table 1 Results of general linear model—univariate analysis for differences in germination capacity (GC), mean germination time (MGT), radicle length (RL), hypocotyl length (HL), fresh weigh of radicle (RFW) and hypocotyl (HFW) and dry weight of radicle (RDW) and hypocotyl (HDR) among species and acid solution treatments
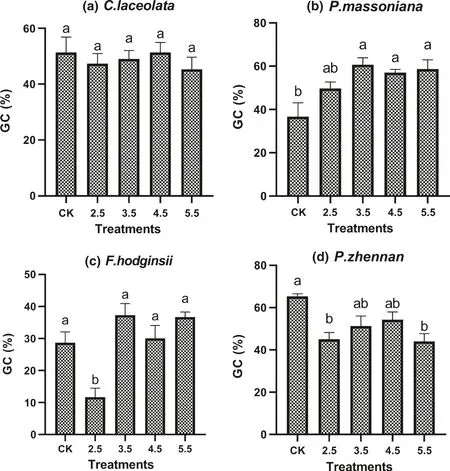
Fig.1 Germination capacity (GC) of seeds of four tree species as affected by simulated acid rain treatments and the control. Values are mean ± SE, and bars followed by different letter are significantly different ( P < 0.05) among treatments
In all simulated acid rain treatments, most of the germination occurred between the 6th and 10th day after sowing. However, the rate of germination, as determined by mean germination time, differed significantly between acid rain treatments and species, but no significant interaction effect was observed (Table 1). The coniferous species ( C. lanceolata and P. massoniana) germinated quickly, about 6 d on average than the broadleaved species for which germination took 12 and 16 d on average for P. zhennan and F. hodginisi, respectively (Table 2). Among simulated acid rain treatments, acid rain treatments with pH 2.5 and 3.5 resulted in quicker germination than that of the control (Table 2).
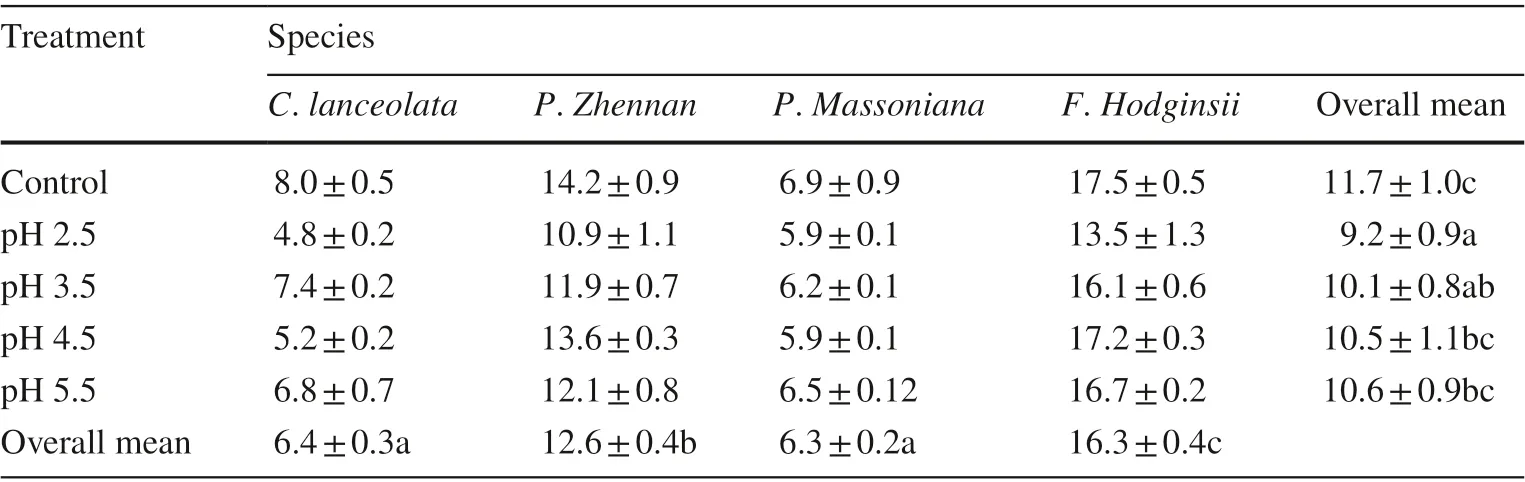
Table 2 Mean germination time (days) of seeds of four south China tree species in response to different acid rain treatments and the control
Seedling emergence
Radicle and hypocotyl length varied significantly among simulated acid rain treatments, species and their interaction (Table 1). Radicle elongation was hindered by acid rain treatment with pH 2.5 compared to other treatments and the control, whereas radicle length was very short in P. massoniana, followed by F. hodginisi and the longest being in C. lanceolata and P. zhennan. At individual species level, radicle elongation of C. lanceolata was reduced by acid treatments of pH 2.5 and 4.5 compared to pH 3.5 (Fig.2 a); that of F. hodginisi was reduced by acid rain treatment of pH 2.5 (Fig.2 b); that of P. massoniana was reduced by acid rain treatment of pH 2.5 compared to pH 3.5 and 4.5 (Fig.2 c); and that of P. zhennan was reduced by pH 2.5 and 3.5 compared to the control while pH 4.5 and 5.5 resulted in statistically similar radicle length to that of the control (Fig.2 d). Hypocotyl length was significantly shorter in acid rain treatment of pH 2.5 compared to other treatments and the control; and the shortest being for P. zhennan followed by C. lanceolata and F. hodginisi and the longest being for P. massoniana. All acid rain treatments reduced hypocotyl elongation of C. lanceolata as compared to the control (Fig.2 e); acid rain treatment of pH 2.5 reduced hypocotyl elongation of F. hodginisi compared to other treatments and the control (Fig.2 f); acid rain treatment with pH 3.5 stimulated hypocotyl elongation in P. massoniana compared to pH 2.5 and the control treatments (Fig.2 g); and acid rain treatments with pH 5.5 stimulated hypocotyl elongation in P. zhennan compared to acid rain treatment of pH 2.5 (Fig.2 h).
Radicle fresh weight varied significantly among species and interaction between species and simulated acid rain treatments while hypocotyl fresh weight varied significantly among species only (Table 1). Averaged overall simulated acid rain treatments; radicle fresh weight was lowest in P. massoniana and F. hodginisi and highest in P. zhennan, whereas hypocotyl fresh weight was higher for P. zhennan than other species. At individual species level, no significant difference in radicle fresh weight was observed for C. lanceolata (Fig.3 a) and F. hodginisi (Fig.3 b), but all simulated acid rain treatments increased radicle fresh weight of P. massoniana as compared to the control (Fig.3 c) while acid rain treatments with pH 2.5 and 3.5 reduced radicle fresh weight of P. zhennan (Fig.3 d). On the contrary, simulated acid rain treatments of pH 2.5 and 5.5 reduced the hypocotyl fresh weight of C. lanceolata compared to acid rain treatment with pH 3.5 and the control (Fig.3 e). For F. hodginisi, hypocotyl fresh weight was reduced by simulated acid rain treatment of pH 2.5 compared to other treatments and the control (Fig.3 f); and it was reduced by acid treatment of pH 2.5 and the control for P. massoniana compared to other acid rain treatments (Fig.3 c) while all treatments had no effect on hypocotyl fresh weight of P. zhennan (Fig.3 h).
Radicle dry weight varied significantly among treatments, species and their interaction, whereas hypocotyl dry weight varied significantly among species only (Table 1). Simulated acid rain treatments of pH 2.5, 3.5 and 5.5 reduced radicle dry weight compared to the control while radicle dry weight of species decreased in the following order: P. massoniana < F. hodginisi < C. lanceolata < P. zhennan. The hypocotyl dry weight was higher for P. zhennan than the other species. Examining at individual species level revealed that radicle dry weight of C. lanceolata was significantly reduced by acid rain treatments of pH 2.5 and 5.5 as compared to pH 3.5 and 4.5 (Fig.4 a); and acid treatment of pH 2.5 and the control resulted in low radicle fresh weight in F. hodginisi than acid treatments with pH 4.5 and 5.5 (Fig.4 b). While all acid treatments had no impact on radicle dry weight of P. massoniana (Fig.4 c), all acid rain treatments reduced radicle dry weight of P. zhennan compared to the control (Fig.4 d). All acid rain treatments significantly reduced hypocotyl dry weight of C. lanceolata compared to the control, particularly the effect was strong with acid rain of pH 5.5 (Fig.4 e). Strongly acid rain (pH = 2.5) significantly reduced hypocotyl dry weight of F. hodginisi and acid rain of pH 4.5 increased hypocotyl dry weight compared to the control (Fig.4 f). For P. massoniana, acid rain treatments of pH 3.5 and 5.5 resulted in higher hypocotyl dry weight than the control and acid treatment of pH 2.5 (Fig.4 g); whereas, none of the treatments had an effect on hypocotyl dry weight of P. zhennan (Fig.4 h).
Discussion
The results provide evidence that the germination response to acid rain depends on the degree of acidity and varies between species. Generally, as the acidity of the rain water increases, the adverse effect on germination increases. However, the adverse effect of acid rain varies between species; with no or stimulatory effect on seeds of conifer species ( C. lanceolata and P. massoniana) and inhibitory effect on broadleaved species ( F. hodginisi and P. zhennan). Generally, production of reactive oxygen species (ROS) is enhanced following environmental stress, such as acid rain. This increased ROS production, in turn, induces oxidative stress, leading to programmed cell death (Kochian et al. 2004). However, species tolerant to such environmental stress activate a suite of antioxidant mechanisms to scavenge the elicited ROS (Foyer and Noctor 2005). The fact that simulated acid rain didn’t affect the germination of C. lanceolata in our study could be related to its high degree of adaptation to acidic environment as it grows naturally in acidic soil in southern China. The stimulatory effect of acid rain on germination of P. massoniana seeds could be related to the relatively thick seed covering that benefited
from scarification effect of the acid treatment. It is worth to note that acid treatments are often used to improve the germination of seeds with thick covering (Tigabu and Odén 2001; Zida et al. 2005). In addition, the hydrogen ion from the acid binds with molecules; thereby reducing their availability for metabolic processes that produces energy (ATP) for embryo growth (Bewley et al. 2013). It should be noted that seed germination is a triphasic process that starts with imbibition of water, the lag phase (a stage where metabolic processes are initiated) and culminated with protrusion of the radicle (Eldhuset et al. 1994; Hartmann et al. 2002). The adverse effect of acid rain with low pH on seed germination has been observed in several species (Lee and Weber 1979; Raynal et al. 1982; McColl and Johnson 1983; Moore and Gillette 1988; Kim 1987; Fan and Wang 2000). Fan and Wang ( 2000) also observed an inhibition of seed germination in three out of five species treated with acid rain of pH 2.0.
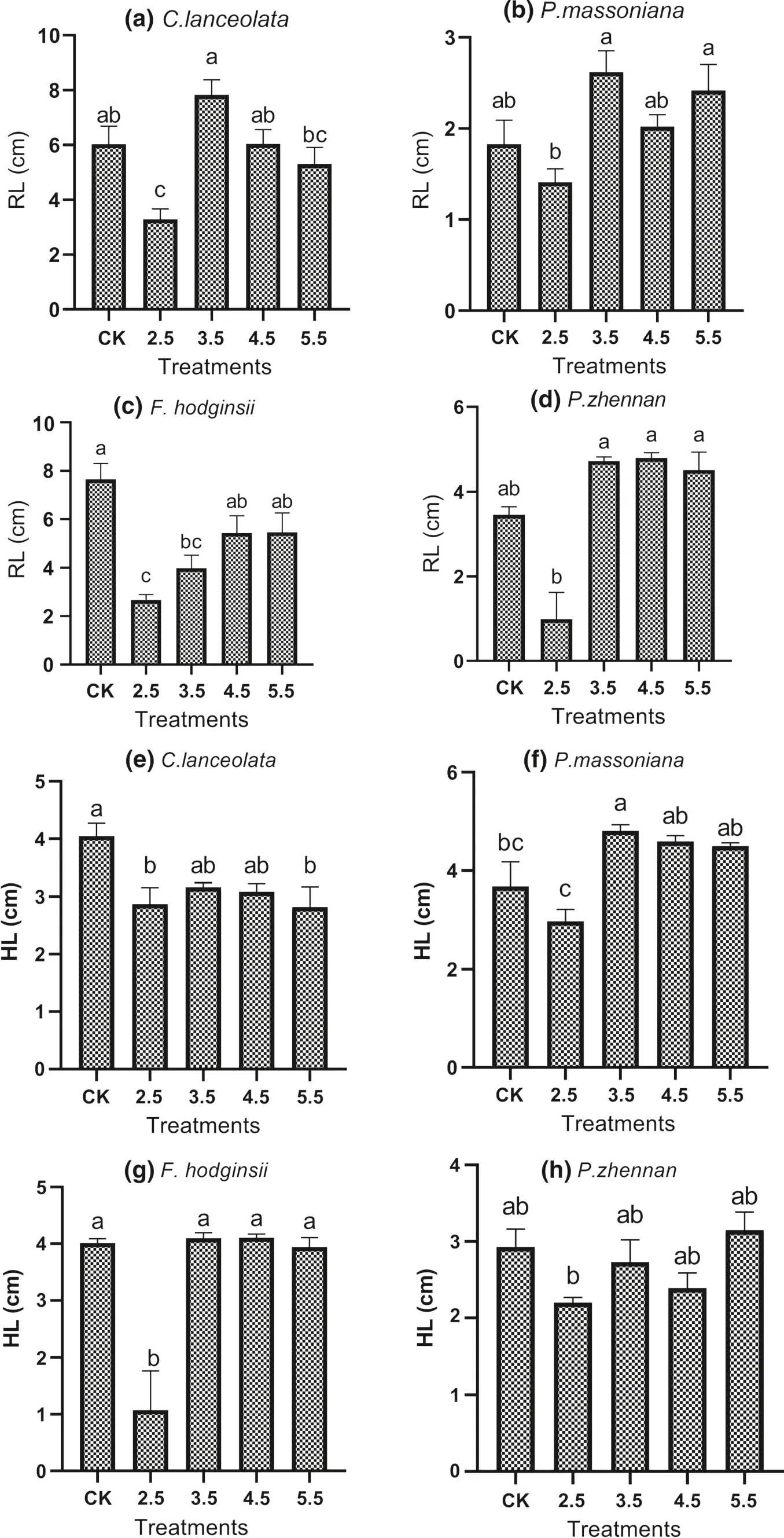
Fig.2 Radicle (RL; a– d) and hypocotyl (HL; e– h) length of seedlings of four tree species as affected by simulated acid rain treatments and the control. Values are mean ± SE, and bars followed by different letter are significantly different ( P < 0.05) among treatments
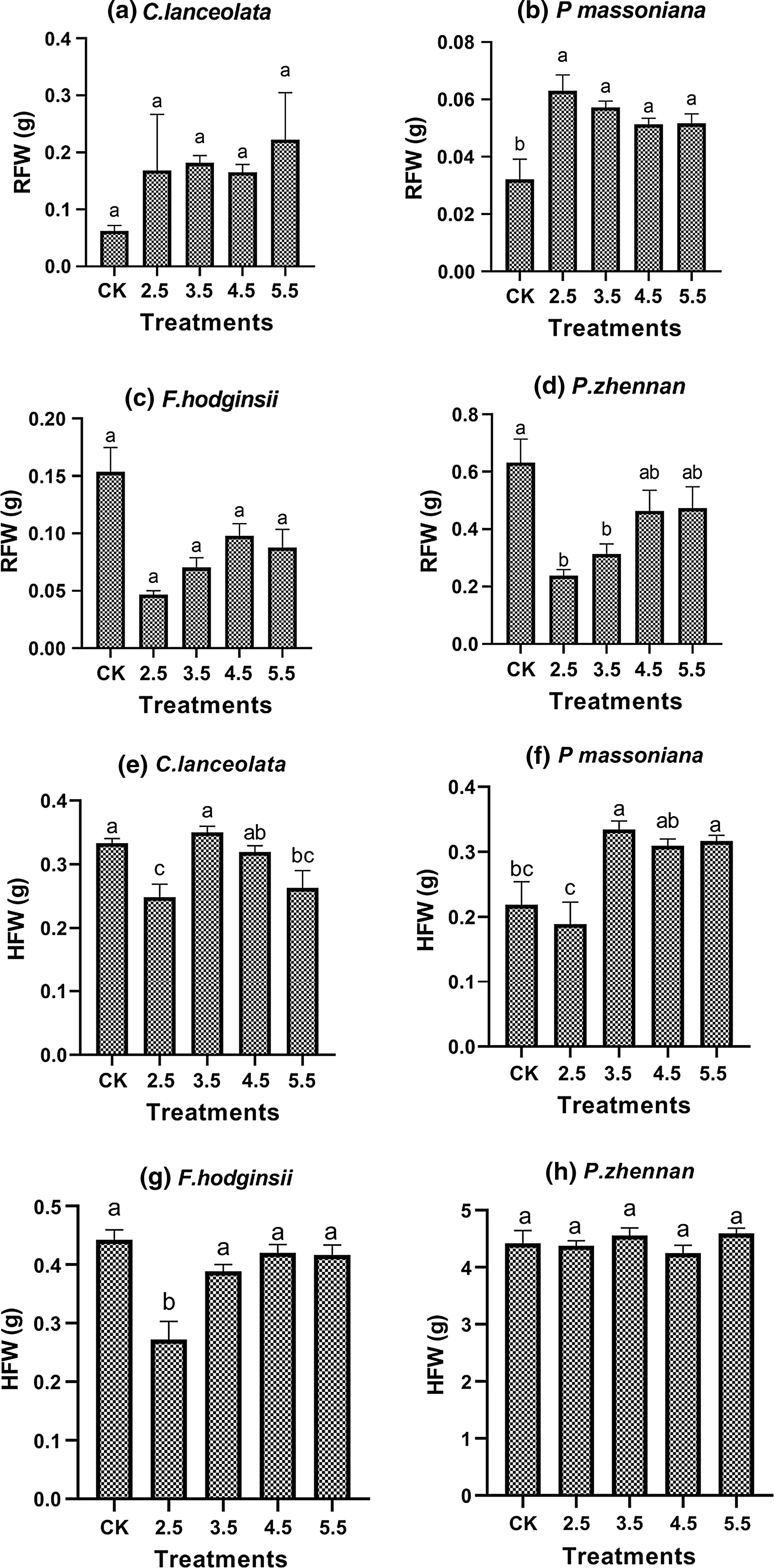
Fig.3 Radicle (RFW; a– d) and hypocotyl (HFW; e– h) fresh weight of seedlings of four tree species as affected by simulated acid rain treatments and the control. Values are mean ± SE, and bars followed by different letter are significantly different ( P < 0.05) among treatments
Unlike seed germination, seedling emergence of all species was affected by acid rain treatments. Radicle and hypocotyl length as well as fresh and dry weight of radicle and hypocotyl were adversely affected by acid rain treatment of pH 2.5. We observed foliar symptoms only on seedlings treated with acid rain of pH 2.5 and lesions occurred on the primary leaves of the species studied. The decrease in seedling growth in response to simulated acid rain of pH 2.0 could be attributed to increased respiratory rate, damage to fine roots and increased leaching of cations (Shan and Feng 1989; Liao and Chen 1992). Plants exposed to acid stress produce a large number of ROS, which denatures membrane lipids, proteins and biological macromolecules, thereby enhancing membrane permeability and lipid peroxidation of cell membrane (Paolacci et al. 2018). Some studies have also found that acid stress can damage organelles such as chloroplasts, mitochondria and nuclei in vivo, which lead to the destruction of cell walls and cell membranes, the deformation of chloroplasts and mitochondria, the disorder of thylakoid arrangement and the disappearance of internal cristae (Lan et al. 2019; Chen et al. 2017). Generally, there is a notion that the response of seed germination to simulated acid rain is not sensitive as compared to subsequent seedling development during early post-germination stages (Raynal et al. 1982). This was supported by several studies that demonstrated that germination was less sensitive to acid rain than seedling establishment (Lee and Weber 1979; Percy 1986; Scherbatskoy et al. 1987). Our results are consistent with the undesirable effects of highly acidic rain water on growth of three legumes (Ashenden and Bell 1989) and other species subjected to acid rain (Halman et al. 2008; Dolatabadian et al. 2013). A study by Park and Yanai ( 2009) also showed that sugar maple seedlings were more sensitive than beech when impacted by acid rain.
The adverse effects of simulated acid rain on seedling development of the studied species might be also exacerbated by the differing anions’ ratio used in this study (i.e., high sulfate to nitrate ratio which was 10:1). Studies have shown that both degree of acidity (pH level) and anion content of simulated rain have a differential impact on seedling development. For instance, high acidity combined with high sulfate in simulated acid rain adversely affected seedling growth in five hardwood species (Fan and Wang 2000). Similarly, high acidity combined with high sulfate decreased volume whereas high acidity combined with high nitrate increased volume of above-ground parts of red spruce (Jacobsen et al. 1990). Apparently, both the degree of acidity of the rain itself and sulfate to nitrate ratio in the acid rain regulates the effects of acid rain on seedling development. In reality, the sulfate to nitrate ratio is high in acid rain in south China; however, further study may be needed to understand if different sulfate to nitrate ratios of simulated acid rain have an impact on the establishment and growth of forest trees. As a whole, broadleaved species appeared to be more sensitive to acid rain than coniferous species, as also observed previously by Evans ( 1982) and Cox ( 1983) based on visible foliar effects and in vitro pollen assays, respectively.
Conclusion
The results demonstrate that germination of seeds and subsequent seedling development of four tree species of south China appeared to be less sensitive to simulated acid rain above pH 3.5. This level of acid rain occurs quite often in South China; thus less acidic precipitation with pH ≥ 3.5 will have little effect on the establishment of the species. The response to acid rain also varies between species; with broadleaved species are more sensitive than coniferous species. Thus, in an environment with high risk of acid rain (pH < 3.5), it would be advisable to plant coniferous species if no acid correction measures are made. Otherwise, acid correction measures, such as liming, would be required to ensure establishment of broadleaved species.
References
Allen HL (1994) Nutritional response of loblolly pine exposed to ozone and simulated acid rain. Can J For Res 24:253–462
Anitha PC, Ramanujan MP (1992) Impact of simulated acid rain on germination and seedling growth of ground nut. Adv Plant Sci 5:180–186
Ashenden TW, Bell SA (1989) Growth responses of three legume species exposed to simulated acid rain. Environ Pollut 62:21–29
Bewley JD, Bradford KJ, Hilhorst HWM, Nonogaki H (2013) Seeds: physiology of development, germination and dormancy, 3rd edn. Springer, New York
Bian YM, Yu SW (1992) Forest decline in Nanshan, China. For Ecol Manag 51(1–3):53
Britton KO, Berrang P, Mavity E (1996) Effects of pretreatment with simulated acid rain on the severity of dogwood anthracnose. Plant Dis 80(6):646–649
Byres DP, Dean TJ, Johnson JD (1992) Long-term effects ofozone and simulated acid rain on the foliage dynamics of slash pine ( Pinus elliottii var. elliottii Engelm.). New Phytol 120(1):61–67
Cao YZ, Wang S, Zhang G, Luo J, Lu S (2009) Chemical characteristics of wet precipitation at an urban site of Guangzhou, South China. Atmos Res 94(3):462–469
Chao WL (1995) Effect of acid rain on the nitrifying activity of copper amended soil left in applied microbial. Lett Appl Microbiol 21:23–24
Chen L, Heerink N, van den Berg M (2006) Energy consumption in rural China: a household model for three villages in Jiangxi Province. Ecol Econ 58(2):407–420
Chen Q, Chen H, Wang W, Liu J, Zhang J (2017) Glycyrrhetic acid, but not glycyrrhizic acid, strengthened entecavir activity by promoting its subcellular distribution in the liver via effl ux inhibition. Eur J Pharm Sci 106:313–327
Cheng Y, Liu Y, Huo M, Sun Q, Wang H, Chen Z, Bai Y (2011) Chemical characteristics of precipitation at Nanping Mangdang Mountain in eastern China during spring. J Environ Sci 23(8):1350–1358
Cox RM (1983) Sensitivity of forest plant reproduction to long range transported air pollutants: in vitro sensitivity of pollen to simulated acid rain. New Phytol 95:269–276
Cui J, Zhou J, Peng Y, He Y, Yang H, Mao J (2014) Atmospheric wet deposition of nitrogen and sulfur to a typical red soil agroecosystem in Southeast China during the ten-year monsoon seasons (2003–2012). Atmos Environ 82:121–129
Dolatabadian A, Sanavy SAMM, Gholamhoseini M, Joghan AK, Majdi M, Kashkooli AB (2013) The role of calcium in improving photosynthesis and related physiological and biochemical attributes of spring wheat subjected to simulated acid rain. Physiol Mol Biol Plants 19(2):189–198
Dong L, Dong H, Fujita T, Geng Y, Fujii M (2015) Cost-effectiveness analysis of China’s Sulfur dioxide control strategy at the regional level: regional disparity, inequity, and future challenges. J Clean Prod 90:345–359
Eldhuset TD, Teigen O, Bjor K (1994) Germination and seedling development. In: Abrahamsen G, Stuanes AO, Tveite B (eds) Longterm experiments with acid rain in norweigian forest ecosystems. Springer, New York, pp 287–294
Evans LS (1982) Biological effects of acidity in precipitation on vegetation: a review. Environ Exp Bot 22:155–160
Fan BT (1993) Environmental chemistry. Zhejiang University Press, Hongzhou (in Chinese)
Fan HB, Wang YH (2000) Effects of simulated acid rain on germination, foliar damage, chlorophyll contents and seedling growth of five hardwood species growing in China. For Ecol Manag 126(3):321–329
Flegal AR, Gallon C, Ganguli PM, Conaway CH (2013) All the lead in China. Crit Rev Environ Sci Technol 43(17):1869–1944
Foyer CH, Noctor G (2005) Oxidant and antioxidant signaling in plants: a reevaluation of the concept ofoxidative stress in a physiological context. Plant Cell Environ 28:1056–1071
Halman JM, Schaberg PG, Hawley GJ, Eagar C (2008) Calcium addition at the Hubbard Brook Experimental Forest increases sugar storage, antioxidant activity and cold tolerance in native red spruce ( Picea rubens). Tree Physiol 28(6):855–862
Hartmann HT, Kester DE, Davies FT, Geneve RL (2002) Plant propagation: principles and practices, 7th edn. Prentice-Hall, Upper Saddle River
Huang K, Zhuang G, Xu C, Wang Y, Tang A (2008) The chemistry of the severe acidic precipitation in Shanghai, China. Atmos Res 89(1–2):149–160
Huang L, Yang J, Zhang G (2012) Chemistry and source identification of wet precipitation in a rural watershed of subtropical China. Chin J Geochem 31(4):347–354
Jacobsen JS, Bethard T, Heller LI, Lassoie JP (1990) Response of Picea rubens seedlings to intermittent mist varying in acidity, and in concentrations of sulfur-, and nitrogen-containing pollutants. Physiol Plant 78:595–601
Kim GT (1987) Effects of simulated acid rain on growth and physiological characteristics of Ginkgo biloba L. seedlings and on chemical properties of the tested soil I. Seed germination and growth. J Korean For Soc 76:99–108
Kochian LV, Hoekenga OA, Pi?eros MA (2004) How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous effi ciency. Annu Rev Plant Biol 55:459–493
Lan X, Yan Y, Yang B, Li X, Xu F (2019) Subcellular distribution of cadmium in a novel potential aquatic hyperaccumulator— Microsorum pteropus. Environ Pollut 248:1020–1027
Larssen T, Schnoor JL, Seip HM, Dawei Z (2000) Evaluation of different approaches for modeling effects of acid rain on soils in China. Sci Total Environ 246(2–3):175–193
Lee JJ, Weber DE (1979) The effects of simulated acid rain on seedling emergence and growth ofeleven woody species. For Sci 25:393–398
Liao LP, Chen CY (1992) Relationships between simulated acid rain, soil acidification and root growth of Chinese fir and Schima superba, Chinese. J Ecol 11:23–28 (in Chinese with an English abstract)
Macaulay BM, Enahoro GE (2015) Effects of simulated acid rain on the morphology, phenology and dry biomass of a local variety of maize (Suwan-1) in Southwestern Nigeria. Environ Monit Assess 187(10):622. https://doi.org/10.1007/s1066 1-015-4844-4
McColl JG, Johnson R (1983) Effects of simulated acid rain on germination and early growth of Douglas-fir and ponderosa pine. Plant Soil 74:125–129
Mei XY, Yang Y, Fang JD (2010) Regime shift of acid rain type in area of Shanghai. Resour Environ Yangtze Basin 19:1075–1079 (in Chinese)
Moore AM, Gillette A (1988) Germination of red spruce and Fraser fir seeds exposed to simulated acid rain. J Elisha Mitchell Sci Soc 104:137–140
Niu YW, Gu JQ, Yu XM, Jiang HR, Dai X (2010) The chemical feature of precipitation in the Yangtze River Delta background region. Environ Chem 29:358–362
Paolacci S, Harrison S, Jansen MA (2018) Are alien species necessarily stress sensitive? A case study on Lemna minuta and Lemna minor. Flora 249:31–39
Park BB, Yanai RD (2009) Nutrient concentrations in roots, leaves and wood of seedling and mature sugar maple and American beech at two contrasting sites. For Ecol Manag 258:1153–1160
Percy K (1986) The effects of simulated acid rain on germinative capacity, growth and morphology of forest tree seedlings. New Phytol 104:473–484
Rajjou L, Duval M, Gallardo K, Catusse J, Bally J, Job C, Job D (2012) Seed germination and vigor. Annu Rev Plant Biol 63:507–533
Raynal DJ, Roman JR, Eichenlaub WM (1982) Response of tree seedlings to acid precipitation: effect of substrate acidity on seed germination. Environ Exp Bot 22(3):377–383
Scherbatskoy T, Klein RM, Badger GJ (1987) Germination responses of forest tree seed to acidity and metal ions. Environ Exp Bot 27:157–164
Shan YF, Feng ZW (1989) Effects of simulated acid rain on seedlings of Cyclobalano glauca Oerst. In: Yang HX (ed) Acid rain and agriculture. China Forestry Press, Beijing, pp 174–178 (in Chinese)
Shi CE, Xueliang D, Yuanjian Y, Xiangrong H, Biwen W (2014) Precipitation chemistry and corresponding transport patterns of influencing air masses at Huangshan Mountain in East China. Adv Atmos Sci 05:1157–1166
Tigabu M, Odén PC (2001) Effects of scarification, gibberellic acid and temperature on seed germination of two multipurpose Albizia species from Ethiopia. Seed Sci Technol 29(1):11–20
Wang H, Han G (2011) Chemical composition of rainwater and anthropogenic influences in Chengdu, Southwest China. Atmos Res 99(2):190–196
Wang WX, Xu PJ (2009) Research progress in precipitation chemistry in China. Prog Chem 21:266–281 (in Chinese)
Xie Z, Du Y, Zeng Y, Li Y, Wu J, Jiao S (2008) Impact of spatiotemporal variation of precipitation on severe acid rain in southern China. Acta Geogr Sin 63:913–923
Zar J (1996) Biostatistical analysis, 3rd edn. Prentice-Hall, Upper Saddle River
Zhang X, Chai F, Wang S, Sun X, Han M (2010) Research progress of acid precipitation in China. Res Environ Sci 23(5):527–532
Zida D, Tigabu M, Sawadogo L, Odén PC (2005) Germination requirements of seeds of four woody species from the Sudanian savanna in Burkina Faso, West Africa. Seed Sci Technol 33(3):581–593
 Journal of Forestry Research2021年2期
Journal of Forestry Research2021年2期
- Journal of Forestry Research的其它文章
- Stem taper functions for Betula platyphylla in the Daxing’an Mountains, northeast China
- Leaf phenotypic variation of endangered plant Tetracentron sinense Oliv. and influence of geographical and climatic factors
- Ecological variations of woody species along an altitudinal gradient in the Qinling Mountains of Central China: area-based versus mass-based expression of leaf traits
- Variations in stem radii of Larix principis-rupprechtii to environmental factors at two slope locations in the Liupan Mountains, northwest China
- A review of ecological mechanisms for management practices of protective forests
- Variation of basic density, calorific value and volumetric shrinkage within tree height and tree age of Ugandan grown Eucalyptus grandis wood