
Seed, germination, and seed-reserve traits diff er along an altitudinal gradient
2022-11-04 09:23:36MuhammetTonguSercannderNesrinlcemalFatihTongu
Journal of Forestry Research 2022年6期
Muhammet Tongu? · Sercan ?nder ·Nesrin Gülcemal · Fatih Tongu?
Abstract Turkish red pine ( Pinus brutia Ten.), an important pine species for aff orestation studies, grows at a wide range of altitudes in the eastern Mediterranean area. Seeds from low (0-400 m a.s.l.), mid (400-800 m) and high(800-1000 m) altitudinal gradients were collected to assess the eff ects of these gradients on morphological traits (seed length, seed width, 100-seed weight), germination variables(germination percentage and mean germination time), and 10 biochemical variables related to seed reserves. Relationships among these variables were examined with a correlation analysis. Seeds from low and high altitudes had signif icantly higher 100-seed weight, higher percentage germination but longer germination times and higher total oil and reducing sugars contents. Seeds from mid altitudes had higher carotenoid and soluble phenolics contents. Among the examined biochemical variables, only f lavonoid content did not diff er among the altitude gradients. The main fatty acids in seeds samples (linoleic, oleic, palmitic and stearic acids) were also inf luenced by the altitude gradients. Oil and total reducing sugars contents were positively correlated with germination percentage, and carotenoid content was negatively correlated with germination percentage. Our results conf irmed that seed and germination variables, including the reserve composition varied signif icantly among the altitudinal gradients.
Keywords Elevation · Germination · Pine · Seed · Seed reserves
Introduction
Turkish red pine (Pinus brutiaTen.), an important forest tree species, is well adapted to mediterranean-type climates and abundant in the eastern Mediterranean area.P. brutiahas become increasingly important for reforestation, timber production and soil stabilization (Tilki and Dirik 2007). The rapid development ofP. brutiaseedlings and their suitability for aff orestation provide very important contributions to wood production. It is also well adapted to diff erent environments, is drought tolerant, and grows naturally in the Mediterranean region from sea level to 1400 m a.s.l., depending on the conditions. In addition, it forms sparse populations along coastal areas of the Black Sea to about 600 m elevation from sea level (Boydak et al. 2006).P. brutiacan also vary greatly in form and growth characteristics depending on the altitudinal range (Dangasuk and Panetsos 2004; Tilki and Dirik 2007).
Trees grow and develop under the combined inf luence of altitude, environment and humans (Peichl and Arain 2006;Turfan et al. 2018). Altitude and environmental factors also aff ect tree lifespan and production of primary and secondary metabolites such as oil, sugar, carotenoids, tocopherol, phenolics, f lavonoids and proteins (Bond et al. 2007). There is a strong representative barrier betweenP. brutiapopulations growing at diff erent altitudinal ranges (I??k and Kara 1997;Tilki and Dirik 2007; ?etin and Boydak 2013). Therefore,germination, biochemical composition and other traits ofP.brutiaseeds may diff er among altitudinal ranges, and specif ic seed resources may need to be chosen for a specif ic altitudinal range to ensure reforestation success. Thus, investigations on adaptive mechanisms of seeds and how seed germination and biochemical contents are altered at diff erent altitudinal ranges are of great importance for conservation and regeneration ofP. bruitaecosystems.
Regeneration (from seed to seedling) is a critical period in the life cycle of pine species and often associated with high mortality rates due to adverse environments and genetic variations in the seeds (I??k 1986; Correia et al. 2014). Pine species show signif icant phenotypic variation for growth, seed morphology and seed mass in response to latitudinal/height gradients. In pines, heavier seeds are associated with faster emergence and seedling growth, especially in extreme conditions (Wennstr?m et al. 2002; Parker et al. 2006; Correia et al.
2014). Therefore, seed mass may also impact germination properties. In addition, seed germination characteristics and seed origins can inf luence seedling characteristics (I??k 1986;Parker et al. 2006). Pine populations also diff er signif icantly in germination percentage and time at diff erent altitudinal ranges (I??k 1986; Tilki and Dirik 2007; Wahid and Bounoua 2013). The adaptation and germination characteristics of seeds to environments at diff erent altitudes can also potentially impact the ontogenic patterns that can determine the success of regeneration and seedling establishment (I??k 1986; Donohue et al. 2010; Correia et al. 2014). Therefore, investigating the variation between germination and biochemical content of seeds at diff erent altitudes is important for forest resource management, as it can contribute to a better understanding of regeneration patterns and can guide the selection of forest reproduction material for successful seedling production and reforestation. Consequently, knowledge of seed biochemical content and germination processes could also be important for seed collection and seedling production.
Although the morphological features and germination properties ofP. brutiaseeds are relatively well known, there is a lack of information about the biochemical composition of seeds and its inf luence on germination. Therefore, the objective of the present study was to evaluate the relationships between seed germination and seed biochemical content ofP. brutiaunder the eff ect of altitude.
Materials and methods
Seed material and seed traits
Seeds from diff erent altitudinal ranges were collected in Kumluca Province of Antalya, Turkey in 2017 (Table 1)and stored at 4 °C until used. These sites are used by the Department of Nursery and Seed Production of the General Directorate of Forestry to collectP. bruitaseeds for seed and seedling production. Cones from the diff erent altitudinal ranges were collected in bulk from pine trees in July, dried in the sun, then seeds were extracted. Wings were removed by scrubbing, and the seeds cleaned. Seed samples (1 kg) were provided by the Forest Management Service of Kumluca.
Pine seed development from pollination to maturity takes 2 years, and seeds are disseminated during the third year (Boydak et al. 2006). Therefore, mean rainfall and temperature from 2015 to 2017 and long-term means were obtained for Antalya Province. Meteorological data was obtained from State Meteorological Service (MGM) and presented in Table 2.
Before germination tests, seeds were f loated in distilled water, and only seeds that were full and sunk were used in the study (Dangasuk and Panetsos 2004). The 100 seed-weights were determined using International Seed Testing Association (ISTA 2009) procedures. To evaluate 100 seed-weights, 800 (100 × 8) from each altitudinal range were randomly sampled to measure seed mass(g) using a precision balance, and 100-seed weights were computed. Seed length (mm) and seed width (mm) were determined with an electronic caliper by measuring 300 seeds (100 × 3) from each altitudinal range.
Germination tests
Germination was tested in the laboratory conditions according to ISTA ( 2009) procedures. Seeds were surfacesterilized by soaking the seeds in sodium hypochlorite solution (1%) for 10 min, then rinsed 3 times with distilled water for 5 min/rinse. For the standard germination test, 200 seeds (50 × 4) from each altitudinal range were placed onto moist f ilter papers in a germination cabinet at 20 ± 0.5 °C for 28 days. Seeds were considered to have germinated if the radicle was at least 2 mm long. Germinated seeds were counted every day for 28 days to determine the germination percentage and mean germination time as described by Y?lmaz et al. ( 2013).
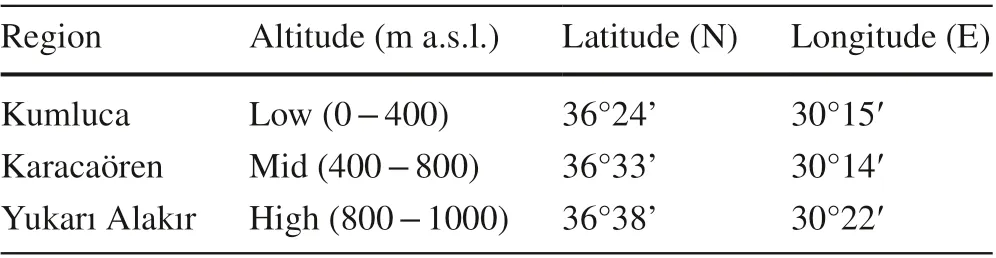
Table 1 Geographic characteristics where seeds of Turkish red pine( P. brutia) were collected in 2017 at diff erent altitudinal ranges in Antalya-Kumluca in Turkey
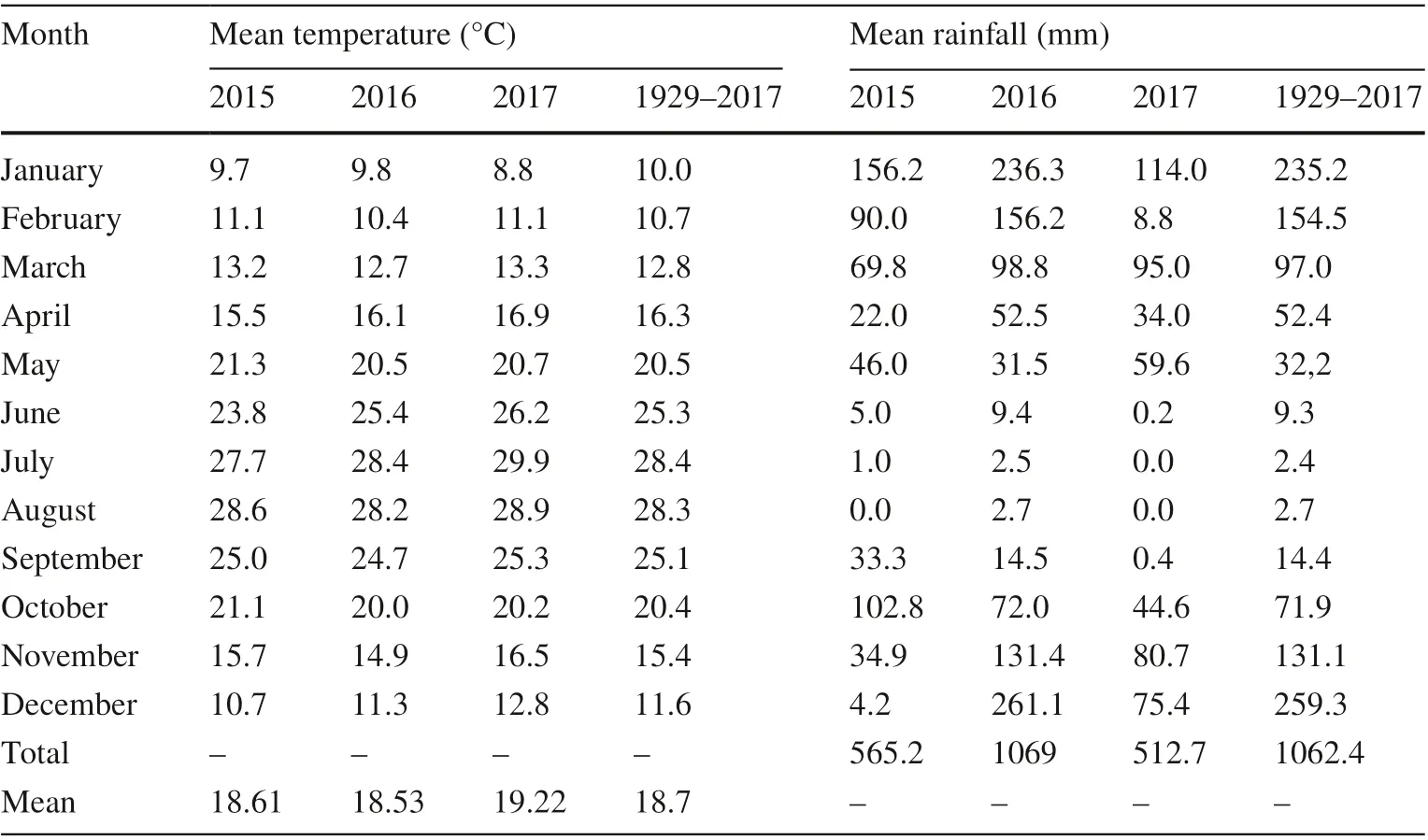
Table 2 Long term(1929 - 2017) and monthly mean temperature (°C) and monthly mean rainfall (mm)for 2015 - 2017 of Antalya province
Biochemical analyses
Seed samples were frozen in liquid nitrogen then ground in a coff ee grinder. For extracting total soluble and reducing sugars, 10 mL of 80% ethanol was added to 1 g of sample, then the samples were shaken vigorously and held at - 20 °C overnight. The next day, samples were centrifuged at 2000 ×gfor 5 min, and the supernatant was transferred to a clean Falcon tube. For measuring total soluble sugars, 1 mL of this supernatant was then combined with 0.5 mL of 5% phenol and 2.5 mL of concentrated sulfuric acid. The samples were kept at room temperature for 10 min to allow color development, then absorbance of the samples was measured at 490 nm (DuBois et al. 1956). For reducing sugars analysis, 2.5 mL of 100 mM sodium borate (pH 9.0)and 0.5 mL of 1% v/v 2-cyanoacetamide was added to 1 mL of supernatant, and the mixture was placed in a boiling water bath for 10 min, then cooled. Absorbance at 280 nm of the samples was measured (Somogyi 1952). A glucose standard was used to obtain the standard curve.
For determining the total amount of carotenoids and xanthophylls, 10 mL of an acetone-hexane solution (4:6) was added to 1 g of sample, and the solution was vortexed for 30 s. Samples were centrifuged at 10.000 rpm for 5 min,then the supernatant was transferred to new tubes and extracted twice with an equal amount of 20% NaCl solution. The upper phases were used to measure absorbance at 436 nm for carotenoids and at 474 nm for xanthophylls.Total carotenoids and xanthophyll contents were calculated according to the methods of the Association of Offi cial Analytical Chemists (AOAC 1984).
The modif ied Emmerie-Engel method was used to determine total tocopherols (Backer et al. 1980) using 0.5 g of sample mixed with 10 mL of acetone-methanol (7:3). The mixture was shaken for 15 min, centrifuged at 10,000 rpm for 20 min, then 1 mL of supernatant was mixed with 1 mL 0.5% w/v 2,2′-bipyridine and 1 mL 0.2% w/v iron chloride.After 60 s, absorbance of the samples at 522 nm was measured. A standard solution of α-tocopherol was used to obtain the standard curve.
For phenolic and f lavonoid measurements, seed samples(0.2 g) were combined with 10 mL of 80% methanol, shaken on a rotary shaker at 120 rpm for 15 min, then centrifuged for 10 min at 4000 rpm. The supernatant was transferred to a new tube, and the pellet was subjected twice to the same extraction procedure just described. The supernatants were pooled, brought to 25 mL with 80% methanol, and used for the following analyses. For total soluble phenolics, the modif ied Folin and Ciocalteu ( 1927) method was used. Folin-Ciocalteu reagent (2 N) and 20% w/v sodium carbonate solution were added to the methanolic extract. The mixture was incubated in the dark for 30 min at room temperature, and absorbance of the samples was measured at 760 nm. Gallic acid was used as the standard. For total f lavonoids using the method of Sakanaka et al. ( 2005), 5% sodium nitrate 10%aluminum chloride and 1 M sodium hydroxide solution were added to the methanolic extract. Absorbance at 510 nm was measured, and a ( +)-catechin standard was used to obtain the standard curve.
For total soluble protein measurements, protein extraction was carried out as described by Tongu? et al. ( 2020).Total soluble protein content was determined by the Lowry-Hartree method (Hartree 1972). 0.90 mL of a solution containing 7 mM potassium-sodium tartrate, 0.81 M sodium carbonate, 0.5 N NaOH solution was added over 1 mL of the protein extract and the samples were incubated in a water bath at 50 °C for 10 min. Subsequently, 0.1 mL of the solution containing 70 mM potassium-sodium tartrate, 40 mM copper sulfate, and 3 mL Folin-Ciocalteu reagent were added to the mixture and incubated for 10 min at 50 °C in a water bath. BSA was used to obtain the standard curve, and absorbance of the samples was measured at 650 nm.
The total oil content of the seeds was measured with nuclear magnetic resonance (NMR) device (Brükermqone),and the results were expressed as a percentage of seed mass.Oil from the ground seed samples was extracted with hexane, the solvent evaporated, and oil samples were converted to fatty acid methyl esters (Marquard 1987). The fatty acid composition was analyzed using gas chromatograph (Shimadzu GC-2025, Japan) equipped with a f lame ionization detector (GC-FID) (Agilent 7890 A, Agilent Technologies Inc., CA, USA) as described by Erba? et al. ( 2016).
Statistical analyses
The germination experiments were performed in an entirely randomized block design, and biochemical analyses were done with three replications. Data were normalized using the arcsine transformation, then subjected to analysis of variance (ANOVA) using SPSS Statistics 22.0 software (IBM,Armonk, NY, USA). Tukey’s test (P≤ 0.05) was used to discriminate the diff erences between the means. To show the relationship between measured parameters, heatmap correlation was calculated using Statgraphics 19 version 19.1.02(Statgraphics Technologies, The Plains, VA, USA).
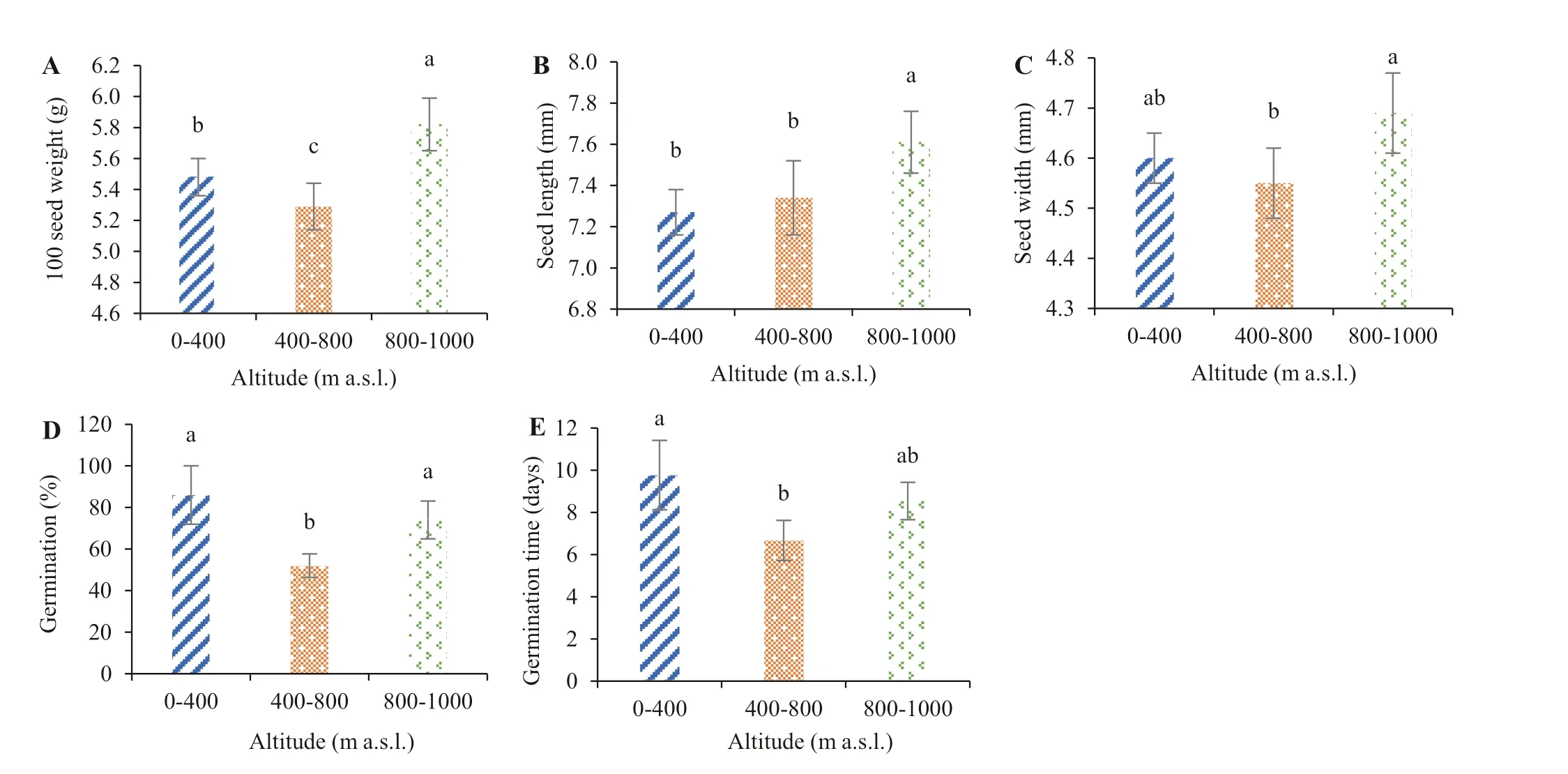
Fig. 1 Mean (± SD) 100 seed weight ( A), seed length ( B), seed width ( C), percentage germination ( D) and mean germination time( E) of P. brutia seeds at three ranges of altitude. Diff erent letters above the bars indicate a signif icant diff erent between the means in Tukey’s test at P ≤ 0.05. The vertical line on the histobars indicate the SD
Results
Seed traits and germination tests of seeds from diff erent altitudinal gradients
There were signif icant diff erences in mean 100-seed weights among the diff erent gradients; 100-seed weight was signif icantly lower at mid altitudes and signif icantly higher at high altitude (Fig. 1 A) compared to low altitude. The lowest 100-seed weight was 5.29 ± 0.15 g for mid altitude, and the highest 100-seed weight was 5.82 ± 0.17 g for high altitude.Seed length increased with altitude, only slightly from low(7.27 ± 0.11 mm) to mid altitude (7.34 ± 0.18 mm), but signif icantly at high altitude (7.61 ± 0.15 mm) (Fig. 1 B). Seed width diff ered among the altitudinal gradients, but the diff erence was signif icant only between the mid and high altitudes(Fig. 1 C).
The diff erent altitudes also signif icantly aff ected germination. Germination percentage of seeds from low, mid and high altitudes was 86 ± 14.05%, 52 ± 5.66% and 74 ± 9.09%,respectively (Fig. 1 D), with only the germination percentage at the mid altitude diff ering signif icantly from the other two. Seeds from the mid altitudes had the shortest mean germination time (6.67 ± 0.95 days), which diff ered signif icantly from the longest mean germination time for seeds at low altitude (9.77 ± 1.65 days) (Fig. 1 E). Mean germination time did not diff er signif icantly between seeds at low and high altitudes.
Biochemical composition of seeds from diff erent altitudes
Total reducing sugars were the highest at the low(8.58 ± 0.39 mg g -1 ) and high (7.89 ± 0.42 mg g -1 ) altitudes, but the difference between the two was not significant, but in comparison the content in seeds from mid altitude was signif icantly lower (5.87 ± 0.55 mg g -1 )(Fig. 2 A). Total soluble sugars content were similar between low (101.07 ± 7.37 mg g-1) and mid altitudes(106.76 ± 3.58 mg g -1 ), but signif icantly lower in seeds from high altitudes (85.41 ± 6.30 mg g -1 ) (Fig. 2 B). Seed samples from the diff erent ranges diff ered in carotenoids and xanthophyll contents. Low- and high-altitude seed samples did not diff er signif icantly in carotenoids, but the content in seeds from mid altitudes (0.12 ± 0.01 μg g -1 ) was signif icantly higher than at the other altitudes (Fig. 2 C).Total xanthophyll content varied signif icantly among the diff erent altitude gradient ranges and xanthophyll content decreased with increased altitude. Xanthophyll content decreased from 0.087 ± 0.00 μg g -1 , to 0.069 ± 0.01 μg g -1 to 0.053 ± 0.00 μg g -1 for low, mid to high altitudes; respectively (Fig. 2 D).

Fig. 2 Mean (± SD) total reducing sugars ( A), total soluble sugars( B), carotenoids ( C), xanthophyll ( D), total soluble phenolics (E),total f lavonoids ( F), tocopherol ( G) and total soluble protein ( H) in P. brutia seeds at three altitudinal ranges. Diff erent letters above the bars indicate a signif icant diff erent between the means in Tukey’s test at P ≤ 0.05. The vertical line on the histobars indicate the SD
The total soluble phenolics increased slightly from low (0.172 ± 0.01 mg g -1 ) to mid altitudes(0.237 ± 0.05 mg g -1 ), but was lowest in seeds from high altitudes (0.124 ± 0.01 mg g -1 ; Fig. 2 E). Total f lavonoid content was not signif icantly diff erent among the altitude ranges, similar to total soluble phenolics content, and was highest (0.609 ± 0.02 mg g -1 ) in seeds from mid altitudes(Fig. 2 F). There was no signif icant diff erences between low (23.81 ± 0.67 mg g -1 ) to mid (24.01 ± 0.04 mg g -1 )altitudes for total tocopherol content, but total tocopherol content was signif icantly higher in seeds from high(29.95 ± 1.27 mg g -1 ) altitudes (Fig. 2 G). Total soluble protein content did not diff er signif icantly between seeds from mid (4.78 ± 0.37 mg g -1 ) and high (4.64 ± 0.11 mg g -1 )altitudes, but content diff ered signif icantly between seeds from low altitudes (5.51 ± 0.27 mg g -1 ) and the other two altitude ranges (Fig. 2 H).
As seen in Table 3, the total oil content and nearly all the fatty acids diff ered signif icantly among seeds from the diff erent altitudinal gradients. The oil fromP. brutiaseeds is rich in unsaturated fatty acids. The major fatty acids were linoleic and oleic acids, making up more than 78% of the total fatty acids in the seeds, followed by palmitic and stearic acids.The fatty acids present in the oils ofP. brutiaseeds were mainly unsaturated fatty acids, making up 83.49%-79.77%of the total, depending on the altitude. Unsaturated palmitoleic and linolenic acids were only present in small quantities. The major saturated fatty acid was palmitic acid, followed by stearic acid. Heptadecanoic acid was only present in trace amounts. Linoleic acid to oleic acid ratio was found to be more than 2:1.
Correlation between seed traits, germination and seed biochemical contents

Table 3 Total oil content and fatty acid composition (%) of P. brutia seeds from the diff erent altitudinal ranges
The result of the correlation analysis for low, mid and high altitudes are given in Fig. 3. Of the 91 Pearson correlation coeffi cients, 25 were signif icant at the 5% level. Among these 25, 12 were positively and 13 were negatively correlated with each other. Germination percentage and mean germination time were each positively correlated with mean germination time (0.95), oil content (0.82) and total reducing sugars (0.88) and negatively correlated with carotenoids content (- 0.80). Seed length was positively correlated with 100-seed weight and tocophereol contents and negatively correlated with total soluble sugars and xanthophyll. Seed width was negatively correlated with total soluble sugars.Oil content, total soluble sugars, carotenoids and xanthophyll contents were positively correlated with total reducing sugars, total soluble phenolics and total soluble proteins;respectively.
Discussion
Our results conf irm the high levels of variation among seed and germination traits found between diff erent altitude gradients ofP. brutiaseeds (I??k 1986; I??k and Kara 1997; Dangusak and Panetsos 2004; Tilki and Dirik 2007; Kurt et al.2012; ?etin and Boydak 2013). High variability for 100-seed weight, seed length and width, germination rate and mean germination times were reported previously forP. brutia(I??k 1986; Dangasuk and Panetsos 2004; Tilki and Dirik 2007; Y?lmaz et al. 2013; Ayan et al. 2019) and for other pine species (Wahid and Bounoua 2013; Correia et al. 2014;Kadri et al. 2015). Seed weight decreases from low altitude to high altitudes, and average 100-seed weight is between 4.0 and 6.7 g inP. brutia(Boydak et al. 2006). I??k ( 1986)reported that 100-seed weight for six populations from two regions of Antalya could be diff erentiated by the growing altitudes. The lowest 100-seed weights were observed from low-altitude populations; mid- and high-elevation populations had higher 100-seed weights. Similarly, altitude was an important factor for 100-seed weight ofP. brutiapopulations in Crete Island; higher altitudes had lower 100-seed weight.In the present study, however, the 100-seed weight was highest at the high altitudes and lowest at the mid altitudes.
Previously reported germination percentages and germination times were higher for low- and mid-altitude populations than for the high-altitude populations ofP. brutia(I??k 1986; Tilki and Dirik 2007; ?etin and Boydak 2013).However, germination percentage and time for seeds from 14P. brutiapopulations, growing from 60 to 925 m a.s.l.,did not exhibit clear diff erentiation amongP. brutiapopulations based on their altitude gradients (Ayan et al. 2019). In the present study, seeds from low- and high-altitude populations had higher germination than the mid-altitude seeds.Mean germination time was lower for mid altitude than low altitude, but the diff erence might be related to the overall lower germination.
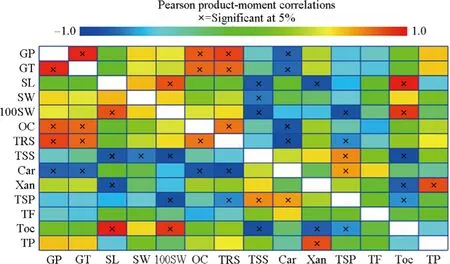
Fig. 3 Relationships and correlation between mean values for 14 seed traits, germination variables, and biochemical components generated by a heat map. Color scale displays the intensity of normalized mean values of diff erent variables. “X” indicates values that were signif icant at 5%. GP germination percentage, GT mean germination time, SL seed length, SW seed width, 100SW 100-seed weight, OC total oil content, TRS total reducing sugars, TSS total soluble sugars,Car carotenoids, Xan xanthophyll, TSP total soluble phenolics, TF,total f lavonoid, Toc tocopherol, TP total soluble protein
Seed characters also diff ered among the altitude gradients forP. brutiain the present study. Seed length and seed width increased with elevation. Previously, mean seed length was found to be 6.6 - 7.9 mm and seed width 3.7 - 4.9 mm, and seed length decreased with increasing altitudes forP. brutia(Boydak et al. 2006). Seed length and width did not diff er signif icantly between seeds from low and mid altitudes, but were signif icantly longer at high altitudes in our study. In contrast, amongP. brutiapopulations in Crete Island, seed length increased from low to mid altitudes but was shorter at high altitudes, and seed width either did not change or was lower at high altitudes (Dangasuk and Panetsos 2004), suggesting that seed morphology is inf luenced not only by altitude but also by the population diff erences and other factors.
Seed length and 100-seed weight decreased with increased altitude inP. brutiain the present study, in contrast to previous reports. Environmental eff ects during the development of seeds, combined with genetic variability could result in variations in seed traits (I??k 1986; Sorensen and Campbell 1993; Wahid and Bounoua 2013). Seed mass is accepted as a criterion of seed quality; seedlings produced from seeds with higher mass tend to have higher germination and growth rates (Parker et al. 2006). Our results showed a correlation between seed mass and germination rate; however, seed mass had a negligible eff ects on seed germination rate forPicea sitchensis(Chaisurisri et al. 1992) andPinus pinasterAit. (Wahid and Bounoua 2013). Therefore, using seed mass as a selection criterion to increase germination rate may not be applicable to all pine species. Seed germination characters may be particularly important for seedling growth (I??k 1986; Wahid and Bounoua 2013), and determining seed traits at diff erent altitudes could help select better seed material. Germination speed appears to have a high heritability (I??k 1986) and be aff ected by reserve composition (Soriano et al. 2011); therefore, seeds collected from appropriate populations and growth conditions may germinate and the resultant seedling grow faster, and survive better when transplanted for aff orestation studies.
The content of sugars, oils and seed proteins, as sources of energy and nitrogen for germination, inf luence the persistence of seeds in the soil and the germination percentage and speed (Soriano et al. 2011; Zhao et al. 2018). The present study revealed high variation in total reducing and soluble sugars, carotenoids, xanthophylls, total soluble phenolics,tocopherol and total soluble protein content inP. brutiaseeds from diff erent altitudinal gradients; only f lavonoid content did not exhibit signif icant changes between altitudes.While many researchers have studied seed size and its inf luence on seedling survival and establishment (Wennstr?m et al. 2002; Parker et al. 2006; Oliver and Borja 2010), seed reserve composition and its relationship to germination has been receiving more attention (Soriano et al. 2011; Wahid and Bounoua 2013). Seed reserve variation is under genetic control and influenced by altitude, and environmental conditions during seed development (Izquierdo et al. 2002;Soriano et al. 2011; Cai et al. 2012).
P. brutiaseeds are rich in oil but the amount of seed oil content was lower in the present study than previously reported (Bagci and Karaagacl? 2004), indicating withinspecies variation, and the total oil content in the seeds is inf luenced by altitude. Seed oil content diff ers among tree species, and oil-rich seeds germinate better and faster and have higher seed mass (Soriano et al. 2011; Kadri et al.2015). In our study, low- and high-attitude seeds had higher oil content, 100-seed weight and germination percentage,conf irming the results of earlier reports; however, germination time was longer for low- and high-altitude seeds than for mid-altitude seeds, which had the lowest oil content.
Sugars are one of the main reserves required during germination and one of the f irst reserves mobilized in germinating seeds (Zhao et al. 2018). Sugar concentration in seeds ofP. brutiais higher than in those of the other pine species(Kadri et al. 2015). Most tree species accumulate sugars as the primary seed reserve, and their accumulation pattern does not vary greatly between years and among populations and is independent from protein and lipid changes (Soriano et al. 2011; Wahid and Bounoua 2013). During germination of seeds ofP. pinasterandP. brutia, sugar concentrations decreased rapidly (Wahid and Bounoua 2013; Tongu? et al.2020). Low- and high-altitude seeds in the present study had higher reducing sugars content and higher germination,but longer mean germination times than seeds from mid altitudes. Based on our results, perhaps sugar concentration aff ects germination rate but not mean germination time.
The total soluble protein content inP. brutiaseeds was lower than the reported values forP. halepensis,P. pinea,P. pinaster, andP. canariensis(Kadri et al. 2015). Seed protein content aff ects germination percentage and speed of tropical deciduous trees andP. pinaster, and protein accumulation in seeds is highly dependent on environmental conditions (Soriano et al. 2011; Wahid and Bounoua 2013). Larger seeds also have lower concentrations of protein and in turn slower germination. Based on protein content of the seeds, seeds of diff erent species have been classif ied as slow or fast germinating (Soriano et al.2011). In the present study, low altitude seeds had the highest protein content (5.51 ± 0.27 mg g -1 ), followed by mid(4.78 ± 0.37 mg g -1 ) and high (4.64 ± 0.11 mg g -1 ) altitude seeds. Though seeds from low altitude had the highest germination rate (86 ± 14.05%), germination rate diff ered signif icantly between high- (74 ± 9.09%) and mid- (52 ± 5.66%)altitude seeds, suggesting that protein content itself is not suffi cient to explain variations in germination rates at different altitudes.
Secondary metabolites in seeds have diverse roles during seed development and germination. Carotenoid compounds that accumulate in seeds take part in abscisic acid formation and regulation of dormancy (Howit and Pogson 2006). Phenolic compounds and tocopherols protect seeds from lipid peroxidation during dormancy and germination (Sano et al.2016). Phenolic compounds also have other roles in seeds such as protection from abiotic stresses, herbivory and insect damage (Mierziak et al. 2014). Phenolic compounds are also involved in dormancy inP. brutiaseeds, and cold moist stratif ication is recommended to break dormancy (Boydak et al. 2006).
Carotenoids are photosynthetic pigments that scavenging reactive oxygen species during stress and act as photoprotective pigments. Carotenoid content ofSolanum kurtzianumvary with altitude (Ibanez et al. 2017).P. brutiaseeds from low and high altitudes had lower carotenoid content and higher germination than in mid-altitude seeds. However, at higher altitudes total xanthophyll content was lower inP.brutiaseeds, which supports the idea that xanthophyll protects the plants against oxidative stress caused by high light intensity (Latowsk et al. 2011).
The total phenolics and f lavonoid contents of pine seeds vary among pine species. Total amount of soluble phenolics in all altitude gradients ofP. brutiawas lower than the other pine species (Kadri et al. 2015). Phenolic compounds protect seeds and their presence induces dormancy and reduce seed germination inP. brutia. I??k ( 1986) observed thatP. brutiaseeds from diff erent altitude gradients at two locations had diff erent germination rates in nursery, but when seeds were washed for 48 h under tap water, there was no diff erence for germination rates of locations and altitude gradients.Similarly, it is recommended that cold moist stratif ication should be applied toP. brutiaseeds to wash off phenolic compounds on seed coat to break dormancy (Boydak et al.2006) and stratif ication improve germination rate ofP. brutiaseeds from diff erent populations (Y?lmaz et al. 2013) and altitude gradients (Tilki and Dirik 2007). In our study, seeds were cold stored, but not pretreated to break any dormancy might be present due to phenolic compounds. The highest total phenolics and carotenoids were found in seeds from mid altitudes. Though the f lavanoid content was not signif icantly diff erent between altitude gradients forP. brutia,mid altitude seed samples had higher f lavonoid content as well. Consequently, mid altitude seed samples exhibited the lowest germination percentage, indicating inhibitory eff ects of phenolic compounds present in seeds is important inP.brutiaseed germination.
Total oil content of seeds was 12 - 14%, and fatty acids detected accounted for more than 97% of the fatty acids present inP. brutiaseeds. There was a clear altitudinal trend inP. brutiaseeds; the amount of saturated fatty acids was higher in high-altitude seeds and the amounts of unsaturated fatty acids decreased with increasing altitude, suggesting that the seeds have some form of altitudinal adaptation that is inf luenced by temperature eff ects (Izquierdo et al. 2002).Depending on the diff erent altitudinal ranges, the ratio of linoleic acid to oleic acid inP. brutiaseeds was more than 2:1, which is consistent with results of studies on lipid prof iles in some pine seeds (Nasri et al. 2004; Kadri et al. 2015).
Seed length but not seed width was signif icantly positively correlated with seed mass. Germination percentage and germination time was also signif icantly positively correlated to each other. Seed mass diff ered among the altitudinal ranges but was not strongly correlated with germination percentage and mean germination time similar to correlation results of I??k ( 1986). Oil content and total reducing sugars showed strong positive correlations to germination percentage and mean germination time, similar to results of Soriano et al. ( 2011). However, soluble protein content did not show signif icant correlations with germination percentage and mean germination time and our results are diff erent from other reports where protein content signif icantly associated with germination variables (Soriano et al. 2011; Wahid and Bounoua 2013). Carotenoids and phenolic compounds have inhibitory eff ects on germination. Total soluble phenolics content had positive correlation with carotenoid content.Although mid altitude seed samples had the highest phenolics and carotenoid content among the altitude gradients,signif icant negative correlations were not detected between germination variables and soluble phenolics content. But carotenoid content had signif icant negative correlations with germination percentage and mean germination time and seed morphological traits, suggesting that germination requirements were at least partially dependent on the content and functioning of phenolics metabolism. Because of the high positive correlation between 100-seed weight and tocopherol, selection ofP. brutiaseeds that have higher tocopherol content may also have longer storability. Our results did not support results reported by Soriano et al. ( 2011) and Wahid and Bounoua ( 2013), who showed positive correlations between germination percentage and protein content.However, we did conf irm the positive correlations for germination percentage, oil and sugars content found by Soriano et al. ( 2011).
The results of the study showed that seed properties, germination percentage and biochemical content ofP. brutiaseeds diff ered signif icantly among the diff erent altitudinal ranges. The germination percentage of seeds in the diff erent altitude gradient range was particularly related to total oil content, indicating that seeds with the highest germination will likely also be rich in total oils. Seeds collected from low altitude had the highest germination, total soluble sugars, xanthophylls and total soluble protein content compared to the others, thus conf irming the superiority of low altitude seeds (I??k 1986). These f indings from diff erent altitudes will provide valuable information for choosing seed properties to select the best seed material and for estimating seed traits and biochemical composition of seeds from diff erent altitudes and designing aff orestation studies and tree improvement programs for diff erent altitudes.
Conclusions
While the study of biochemical mechanisms underpinning altitudinal adaptation of seed reserves in plants is still in its infancy, the available data illustrate that altitude and local environmental conditions can leave strong morphological,physiological and biochemical signatures inP. bruitaforests.Our results conf irmed the diff erent altitudinal ranges signif icantly inf luence seed morphological and germination traits and reserve composition. We also conf irmed the positive relationship between seed size and germination percentage.The germination percentage at diff erent altitudes was related to total oil and reducing sugars contents, but we did not f ind a signif icant positive relationship between germination percentage and protein content. The signif icant negative correlation between germination percentage and carotenoid content suggests that secondary metabolites are also involved in germination performance ofP. brutiaseeds. Changes in total saturated and unsaturated fatty acid contents among altitude gradient ranges showed the inf luence of environment on seed reserve composition. These f indings provide valuable information on seed properties for selection of seed material based on altitude and biochemical content of seeds.
Acknowledgements We thank the forest management service of Kumluca (Antalya) for providingP. brutiaseed samples and information on the locations. Sercan ?nder was f inancially supported by Council of Higher Education under Y?K 100/2000 fellowship program for graduate students.
Author contributions Muhammet Tongu?: designed the study, carried out the experiments, analyzed the data; Sercan ?nder: carried out the experiments, analyzed the data, generated the illustrations; Nesrin Gülcemal: carried out the experiments; Fatih Tongu?: designed the study, carried out the experiments.
 Journal of Forestry Research2022年6期
Journal of Forestry Research2022年6期
- Journal of Forestry Research的其它文章
- Mixing planting proportions in a plantation aff ects functional traits and biomass allocation of Cunninghamia lanceolata and Phoebe bournei seedlings
- Intraspecif ic leaf morphological variation in Quercus dentata Thunb.: a comparison of traditional and geometric morphometric methods, a pilot study
- Mapping the potential distribution suitability of 16 tree species under climate change in northeastern China using Maxent modelling
- Correction to: Eff ects of active molecules of Korean pine seed on rodent health and implications for forest regeneration
- Analysis of the chloroplast genomes of four Pinus species in Northeast China: Insights into hybrid speciation and identif ication of DNA molecular markers
- Genetic variation and superior provenances selection for wood properties of Larix olgensis at four trials