
Clonal isolation of endothelial colony-forming cells from early gestation chorionic villi of human placenta for fetal tissue regeneration
2020-05-07 01:19:20KewaGaoSiqiHePriyadarsiniKumarDianaFarmerJiandaZhouAijunWang
World Journal of Stem Cells 2020年2期
Kewa Gao, Siqi He, Priyadarsini Kumar, Diana Farmer, Jianda Zhou, Aijun Wang
Kewa Gao, Siqi He, Jianda Zhou, Department of Burns and Plastic Surgery, The Third Xiangya Hospital of Central South University, Changsha 410013, Hunan Province, China
Kewa Gao, Siqi He, Priyadarsini Kumar, Diana Farmer, Aijun Wang, Surgical Bioengineering Laboratory, Department of Surgery, University of California Davis, Sacramento, CA 95817,United States
Kewa Gao, Siqi He, Priyadarsini Kumar, Diana Farmer, Aijun Wang, Institute for Pediatric Regenerative Medicine, Shriners Hospitals for Children, Sacramento, CA 95817, United States
Aijun Wang, Department of Biomedical Engineering, University of California Davis, Davis, CA 95817, United States
Abstract
Key words: Placenta; Endothelial colony forming cells; Chorionic villi; Angiogenesis;Tissue engineering
INTRODUCTION
Over the past three decades, fetal medicine, especially fetal surgery, has been substantially developed for the treatment of congenital disorders. These include structural defects, such as spina bifida, congenital diaphragmatic hernia,sacrococcygeal teratoma, cardiac malformations[1-3], and genetic disorders, such as hemophilia[4,5], Duchenne muscular dystrophy[6]and cystic fibrosis[7,8]. For the past several years, our group has been exploring and establishing stem cell-based regenerative fetal treatments combined with tissue engineering for a variety of congenital disorders. For instance, we have successfully isolated placental mesenchymal stem/stromal cells (PMSCs) from the chorionic villus of early gestation placentas, and developed a PMSC-based fetal treatment for spina bifida (SB)[3,4,9-12].Using the surgically-created fetal ovine SB model, we showed that augmentingin uterosurgical repair of SB defects with PMSCs can rescue neurons and cure SBassociated motor function deficits at birth[3,9-11]. However, consistent with numerous other cases in which therapeutic effects were observed using MSCs, the transplanted PMSCs did not persist following transplantation, nor contribute to tissue regeneration by integration[3,13-17]. Rather, the PMSCs rescued neuronsviaparacrine mechanisms. In the aforementioned studies, small intestinal submucosa extracellular matrix (SISECM) was the biomaterial scaffold used to deliver the stem cellsin vivo[9,18,19]. Porcine small intestinal submucosa (SIS) scaffold is a Food and Drug Administrationapproved natural ECM, which serves as a suitable provisional matrix for tissue regeneration. Based on our previous application of SIS-ECM, we believe it can be useful as a scaffold for delivering various types of stem cells to different affected tissues.
In another recent study of hemophilia A, we showed that when co-transplanting PMSCs with cord blood-derived endothelial colony-forming cells (CB-ECFCs), PMSCs integrated into the host environment and formed stable, long-term engraftment[20].This observation suggests that ECFCs play a critical role in facilitating the long-term survival and engraftment of PMSCs[20].
The potential of ECFCs is also noted in vascularization, including vasculogenesis and angiogenesis. Vasculogenesis is thede novoformation of blood vessels, and is an essential physiological process that occurs during embryonic development and tissue regeneration. Angiogenesis is the growth of new capillaries from pre-existing blood vessels, which is observed both prenatally and postnatally[21]. ECFCs are highly proliferative endothelial progenitor cells that can differentiate into mature endothelial cells[22], and facilitate the functional formation of angiogenesis and thus vascularization. Therefore, cell therapies using ECFCs isolated from various tissue sources, such as bone marrow[23], adipose tissue[24], peripheral blood[25]and cord blood[20,26], have been sought as a therapeutic method to improve vascularization for various disorders[27]. Vascularization is vital to the development, maintenance, and regeneration of tissues. Angiogenesis, one vascularization process in which new blood vessels are formed from preexisting ones, plays a crucial role in embryonic and fetal development[21,28]. A defect in angiogenesis can lead to a variety of diseases, such as heart and brain ischemia, neurodegeneration, hypertension, osteoporosis, respiratory distress, and preeclampsia, to name a few[29]. Therefore, improving angiogenesis can ameliorate these aforementioned disorders by substantially increasing the supply of nutrients and oxygen to the affected tissues, and thus subsequently promoting tissue regeneration and functional repair[30-32]. Furthermore, the proliferative capacity of ECFCs, as well as their ability to integrate into the circulatory system, has allowed them to also be used as a delivery method of mutant genes to treat genetic vascular diseases[20,33]. Overall, the potential of ECFCs is greatly noted, and they may be ideal for treating the various disorders listed above, both adult and congenital. For example, an ideal long-term treatment strategy for congenital genetic diseases, such as hemophilia, is to apply appropriate stem cells during the first trimester of gestation,and treat the fetus prior to the development of a fetal immune system[4,34].
The placenta is a highly vascularized organ that plays a pivotal role in supporting and regulating fetal development with active vascularization beginning at an early gestational age[35]. During the first trimester of gestation, the placenta rapidly develops from the trophectoderm. The developmental process includes the formation of the villus tree and the extensive vasculature necessary to support the developing fetus.Hence, the early gestation placenta may pose a source from which we can reliably obtain a variety of progenitor cells such as ECFCs, in addition to the PMSCs that we have already established[35-37].
Several methods have been established to isolate ECFCs from term placentas. Patelet al[38]isolated large numbers of ECFCs by flow cytometry enrichment of CD45-CD34+-CD31Locells from term placentas, and found that placenta-derived ECFCs possessed angiogenic qualities similar to CB-ECFCs. Solomon and colleagues obtained ECFCs from the micro- and macrovascular tissues of human term placentas using CD31 magnetic bead sorting and colony isolation[39]. Thus far, a protocol for isolating ECFCs from early gestational preterm placentas has not been wellestablished, and the feasibility of using these ECFCs as an autologous source for fetal treatment has not been explored. In this study, we established a method for isolating ECFCs from early gestation placentas, and characterized their phenotype and angiogenic functions.
MATERIALS AND METHODS
Isolation of single cells from the chorionic villus of early gestation human placenta by enzymatic dissociation
Discarded human early gestation (GA 12-16 wk) placentas were collected from the University of California, Davis, Medical Center. The study was submitted to the University of California, Davis, Institutional Review Board, and was determined to be exempt from review. The chorionic villi of the placenta were dissected into small pieces and washed three times with 1X phosphate-buffered saline (PBS) solution containing 100 IU/mL of penicillin and 100 μg/mL of streptomycin. Chorionic villus tissue (100 mg) was placed in a 100 mm dish and digested with 10 mL of enzyme solution containing 1 mg/mL collagenase Type I (Gibco), 0.1% trypsin (Invitrogen),and 0.2 mg/mL DNase (Invitrogen) by incubating at 37 °C, 5% CO2for 20 min. The cell suspension was collected, neutralized with a medium containing 10% fetal bovine serum (FBS), and placed on ice. Fresh enzyme solution was added to the remaining tissue in the petri dish and incubated for an additional 20 min. The collection of cells and enzyme digestion was repeated until the whole tissue was completely dissociated. The cell suspensions were pooled, filtered through 70 μm nylon mesh,and incubated with red blood cell lysate buffer for 5 min at room temperature.
Sorting CD31-positive cells by magnetic beads and culture
Cells that were obtained by enzyme dissociation, as described above, were labeled with magnetic bead antibodies, and sorted according to the manufacturer’s protocol(MACS cell separation system; Miltenyi Biotec). Briefly, cells were first labeled with PE-mouse anti-human CD31 antibody and then bound to anti-PE microbeads(MiltenyiBiotec). Labeled cells were passed through a separation column under the magnetic field, and CD31-positive cells were collected and seeded on a rat-tail collagen Type I (BD Biosciences Discovery)-coated tissue culture treated dish at a low seeding density (2000-4000 cells/cm2). Cells were cultured in Endothelial Cell Growth Medium MV-2 media (ECGM-MV2, PromoCell) with the addition of 250 ng/mL TGFβ inhibitor SB431542 (Stemcell Technologies) and 10 ng/mL vascular endothelial growth factor (VEGF; R&D Systems). Cells were fed every day for the first 7 d and every 2 d until the appearance of cell colonies.
Isolation of cell clones using the cloning cylinder
Once a cell colony with a cobblestone-like morphology appeared and grew to about 20 cells, it was hand-picked using a cloning cylinder. The rim of a sterile cloning cylinder (Millipore/Sigma) was coated with sterile vacuum grease and placed onto the location of the target clone marked under the microscope. Trypsin-EDTA was used to detach the cells within the cloning cylinder. All cells obtained from each of the selected colonies was seeded into one well of a 24-well plate (Corning). After about 2-3 wks of culture and upon reaching about 90% confluency, the cells were passaged into a 6-well tissue culture treated dish. In addition, 5000 cells from each colony were seeded into one well of a 96-well plate for CD31 and VE-Cadherin staining to confirm the ECFC phenotype. The monoclonal cells that displayed cobblestone-like morphology and co-expressed CD31 and VE-Cadherin underwent further expansion.CV-ECFCs were frozen at passage five and subsequently used for all experiments.
Cord blood-derived ECFCs isolation and culture
Cord blood was collected from discarded term placentas obtained from the University of California Medical Center. CB-ECFCs were isolated as previously described[20].They were cultured in ECGM-MV2 media (PromeCell). CB-ECFCs between passages 4-6 were used in this study.
Flow cytometry
Flow cytometry was used to characterize the cellular composition of the chorionic villus of human early gestation placental tissue, and the surface markers of isolated and expanded CV-ECFCs. The Attune NxT Flow Cytometer (ThermoFisher Scientific)was used for performing flow cytometry, and FlowJo software (FlowJo LLC) was used for data analyses. All antibodies were obtained from BD Biosciences. For characterization of early gestation placenta cellular composition, single cells obtained from enzymatic dissociation of placental villi were fractionated into tubes containing approximately 5 x 105cells per sample and stained with PE-CD31 (555446), APC-CD13(561698), APC-CD90 (561971), APC-CD34 (560940), APC-CD45 (560973), APC-CD105(562408), APC-CD117 (561118), and Alexa Fluor 647-CD146 (563619). For phenotypic characterization of CV-ECFCs, cells were stained with PE-CD31 (555446), PE-CD34(555446), PE-CD144 (560410), PE-CD90 (561972), PE-CD45 (555483), APC-CD105(562408), Alexa Fluor 647-CD146 (563619), and Alexa Fluor 647-CD309 (560495). PEMs IgG1 κ (555749), APC-Ms IgG1 κ (550854), and Alexa Fluor 647-Ms IgG1 κ (557783)were used as isotype controls, and anti-mouse Igκ CompBeads were used to generate compensation controls. The LIVE/DEAD? Fixable Near-IR Dead Cell Stain Kit(ThermoFisher Scientific) was used to exclude interference from the non-specific staining of dead cells. CV-ECFCs were transduced using an established protocol[20].Transduction efficiency was assessed by tdTomato expression quantified by flow cytometry.
Immunohistochemistry and immunocytochemistry staining
Chorionic villus tissue was dissected from human early gestation placentas, fixed with 10% formalin for 24 h, protected by 30% sucrose dehydration until the tissue specimen settled to the bottom of the tube, and then embedded in O.C.T compound(Sakura Finetek). Serial sections were made at 6 μm thickness using a Cryostat (Leica CM3050S) and mounted onto microscope slides (Matsunami Glass). O.C.T compound was washed off by water. Tissue sections were permeabilized by incubating the tissues with PBS containing 0.5% Triton X-100 for 10 min. Tissue sections were then incubated with PBS containing 5% bovine serum albumin (BSA) for 1 h at room temperature to block non-specific binding sites. Samples were incubated with primary antibody diluted in PBS containing 1% BSA at 4 °C overnight. The dilutions of primary antibodies were: Mouse anti-human CD146 (BD Biosciences) 1:50, mouse anti-human NG2 (BD Biosciences) 1:50, mouse anti-human CD34 (Dako) 1:25, rabbit anti-human von Willebrand factor (vWF) (Dako) 1:200. Tissue sections were then stained with the respective secondary antibodies: donkey anti-rabbit, conjugated with Alexa647 (ThermoFisher Scientific) and donkey anti-mouse, conjugated with Alexa647(ThermoFisher Scientific), diluted 1:500 with 1% BSA in PBS, and incubated for 1 h at room temperature. The slides were then counterstained with 1:5000 dilution of DAPI for 5 min, mounted with Prolong Diamond Antifade Mountant (Invitrogen), and imaged with a Zeiss Observer Z1 microscope. For CV-ECFC immunocytochemical characterization, cells were seeded in 96-well plates and fixed with 10% formalin.Cells were stained using rabbit anti-VE-Cadherin (Cell Signaling) at 1:400 dilution or mouse anti-CD31 antibodies (Dako) at 1:40 dilution and imaged, as described above.
Acetylated low-density lipoprotein uptake
The CV-ECFCs were cultured in 0.5% BSA for 24 h and then incubated with 10 μg/mL Dil-Ac-LDL (Alfa Aesar) in serum-free culture medium at 37 °C for 5 h. Cells were then washed three times with PBS, fixed with 10% formalin for 15 min, and stained with DAPI (1:5000 in water) to label nuclei. The cells were imaged using a Zeiss Observer Z1 microscope.
Tube formation assay
Twenty-four-well culture dishes were coated with 300 μL Matrigel (BD Biosciences)per well and allowed to gel for 60 min at 37 °C. CV-ECFCs (100,000) were seeded onto the Matrigel-coated wells and incubated at 37 °C, 5% CO2. Phase contrast images were acquired 12 h after seeding using a Zeiss Observer Z1 microscope. Tube formation was quantified using the angiogenesis analyzer in ImageJ software for total junction numbers and total branch lengths.
Genetical modification and viral transduction of CV-ECFCs
CV-ECFCs could serve as a promising autologous cell source for the treatment of genetic diseases. To test if CV-ECFCs can be genetically modified, we transduced CVECFCs using lentiviral vectors for proof-of-concept evaluations. All lentiviral constructs were generated at the Institute for Regenerative Cures (IRC) Vector Core,University of California, Davis. CV-ECFCs were transduced in a transduction medium composed of DMEM high glucose, 10% FBS and 8 μg/mL protamine sulfate(MP Biomedicals) with pCCLc-MNDU3-LUC-PGK-Tomato-WPRE for quantitative analysis at a multiplicity of infection (MOI) of 10 for 6 h. After 6 h, the transduction medium was discarded, and cells were cultured in ECGM-MV2 medium for 72 h. A fluorescence microscope (Zeiss Observer Z1) was used to observe tdTomato expression, and flow cytometry was used to detect transduction efficiency.
Cytocompatibility of CV-ECFCs with biomaterial scaffolds
CV-ECFCs represent a promising new cell source for various fetal or adult tissue engineering applications. To test the cytocompatibility of CV-ECFCs with biomaterial scaffolds, we used a clinical grade SIS-ECM scaffold as a representative scaffold to be testedin vitro. Punch-outs (6 mm) of the SIS-ECM scaffold (Biodesign??Dural graft,Cook Biotech, West Lafayette, IN) were incubated in the culture medium overnight. A quantity of 2 × 105transduced CV-ECFCs were suspended in 10 μL culture medium,seeded onto the pre-equilibrated SIS-ECM scaffold, and cultured at 37 °C, 5% CO2for 4 h to allow cells to attach. Then, the culture medium was subsequently added to cover the CV-ECFC/SIS scaffold composite, and incubated at 37 °C, 5% CO2for 24 h.A fluorescence microscope (Zeiss Observer Z1) was then used to observe cell morphology on the scaffold.
Statistical analysis
All data analysis was performed using PRISM 7 software (GraphPad Software Inc.).Descriptive statistical data are reported as mean ± SD.
RESULTS
Cellular composit ion characterization of the human early gestation placenta chorionic villus
The placenta is composed of a variety of progenitor cells, and undergoes rapid development during the early gestational period of pregnancy. Hence, it serves as a unique source for obtaining progenitor cells that could be utilized for tissue engineering purposes. We first analyzed the cellular composition of early gestation placenta villi. The typical endothelial cell markers CD31, CD34 and vWF were used to characterize the endothelial cells present in the chorionic villus tissue (gestation age of 14 wk 5 d). Results confirmed that these endothelial cell markers were broadly distributed throughout the chorionic villus tissues (Figure 1A, the first three panels).CD146 is a marker expressed on the endothelium in capillaries and perivascular cells around the venules. We found that CD146-positive cells were widely present in placental villi tissues (Figure 1A, the 4thpanel). Flow cytometry quantitative analysis was used to characterize the cell phenotypes derived from the enzymatic dissociation of placental tissue. We found that during early gestation (12-16 wk), placental chorionic villus tissues had 52.3% ± 24.1% CD45-positive cells , 43.9% ± 10.39% CD13-positive cells, 23.98% ± 14.28% CD90-positive cells, 24.5% ± 5.47% CD105-positive cells, 23.83% ± 13.66% CD31-positive cells, 14.79% ± 7.65% CD34-positive cells, 9.57%± 2.5% CD146-positive cells, and 7.35% ± 10.79% CD117-positive cells (Figure 1B and C). Since CD31 is an established, specific endothelial cell marker, it was selected for the magnetic sorting strategy of CV-ECFCs from chorionic villi. We also analyzed the co-expression of CD31 and other markers on the isolated cells derived from early gestation placental chorionic villus tissues. We found that 12.56% ± 4.24% of CD31-positive cells were also CD45 positive, and most likely of the non-adherent hematopoietic lineage. A subpopulation of CD31-positive cells (4.22% ± 1.53%) were also positive for CD34, a marker of immature endothelial progenitor cells. This subpopulation will most likely be able to grow into colonies with high proliferative potential. A subpopulation of CD31-positive cells (1.34% ± 0.67%) were also positive for CD90, and CD90 is generally considered as an MSC marker (Figure 1D). Although the cells that simultaneously expressed CD31 and CD90 account for only a small fraction of all cells, their higher proliferative capacity overtook the expansion of CD31-sorted CV-ECFCs. Hence, manual cloning was necessary to obtain pure populations of CV-ECFCs.
Clonal isolation of CV-ECFCs by magnetic beads sorting
By enzymatically dissociating tissues, we obtained 1.22 × 106± 0.32 × 106single cells from 100 mg of chorionic villus tissue (n= 5). Single cells were then subjected to magnetic bead sorting and manual cloning. After 4-6 passages and confirmation of surface markers, an ECFC cell line with proliferative potential was obtained. A brief illustration of this process is shown in Figure 2A. The CD31-positive cells were added to ECGM-MV2 containing TGF-β inhibitor and VEGF. Cell colonies grown from single cells appeared 5-10 d after being cultured, and they presented with multiple morphologies (Figure 2B). CV-ECFC colonies exhibited a cobblestone-like morphology, whereas MSC clones exhibited a spindle-shaped morphology. There were also colonies that exhibited non-uniform morphologies. Cell colonies that displayed a cobblestone-like morphology were chosen using a cloning cylinder,seeded, and cultured in a 24-well plate. In order to get a uniform population and increased cell number for subsequent characterization by immunocytochemistry, cells underwent two additional passages. Those cell lines that co-expressed CD31 and CD144 were expanded for subsequent experiments (Figure 2C). After a total of four passages within 6-8 wk of culture, a total number of 1.8 × 107CV-ECFCs per 100 mg of chorionic villus tissue was obtained.
Flow cytometry analysis of CV-ECFC morphology
The immunophenotypic profile of CV-ECFCs was analyzed by flow cytometry. These cells were positive for well-established ECFC surface markers, including CD31(99.55% ± 0.49%), CD144 (98.95% ± 0.64%), CD146 (98.3% ± 1.98%), CD105 (97.7% ±1.56%), CD309 (39.6% ± 0.99%), and low expression of CD34 (23.2% ± 12.87%). They were negative for hematopoietic and MSC surface markers CD45 (1.87% ± 2.18%) and CD90 (2.95% ± 4.02%) (n= 3) (Figure 3). The expression levels of various surface markers were similar to CB-ECFCs, as previously published[20], except for the expression level of CD34 of CV-ECFCs, which was higher than that of CB-ECFCs(2.16% ± 1.06%).

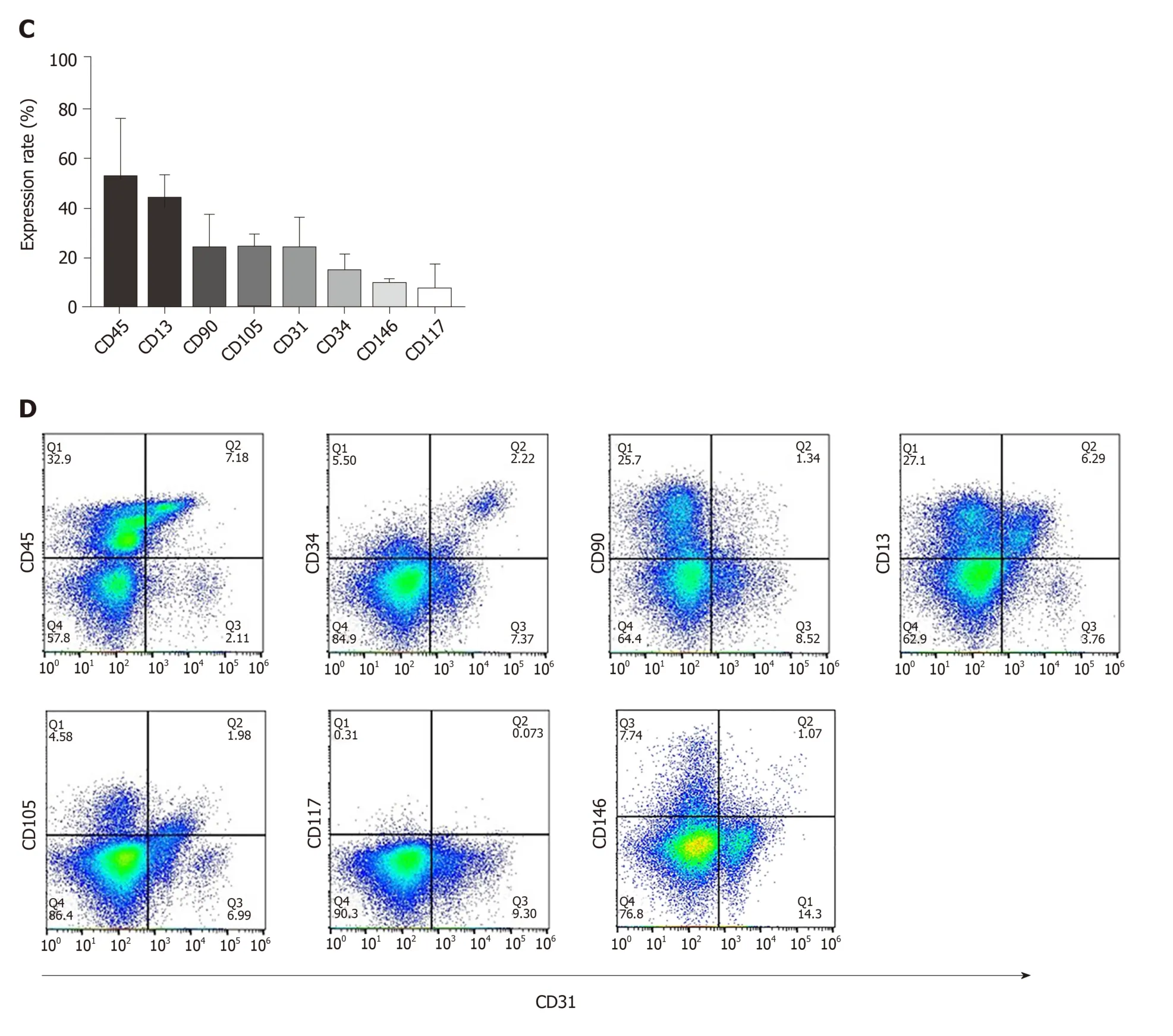
Figure 1 Characterization and cellular composition of the human early gestation placenta. Cellular composition of the human early gestation placenta chorionic villi was characterized. A: Immunofluorescence staining of CD31, CD34, von Willebrand factor and CD146 showed positive cells, depicted with arrows, located in chorionic villi and organized into blood vessel structures; B, C: Flow cytometry analysis of the surface markers of single cells derived by enzymatic dissociation of the placental chorionic villi; each marker’s expression is quantitively presented (data were expressed as mean ± SD, n = 3); D: Flow cytometry analysis of the coexpression of typical endothelial cell marker CD31 and other surface markers. Scale bar = 100 μm.
CV-ECFCs are functionally comparable to CB-ECFCs
Tube formation and Dil-Ac-LDL uptake experiments are commonly used to identify endothelial cells from a functional perspective. Like CB-ECFCs, CV-ECFCs formed a similar tubular structure on the surface of Matrigel, which can persist for more than 48 h and then disintegrate. The capacity ofin vitroangiogenesis of CV-ECFCs and CBECFCs showed no significant difference in quantity by the angiogenesis analyzer in ImageJ (Figure 4A). These CV-ECFCs showed a comparable ability of Dil-Ac-LDL uptake to CB-ECFCs (Figure 4B).
To explore the possibility of CV-ECFCs expressing exogenous proteins, and their potential for treating genetic defects, the transduction rate of these cells was explored.CV-ECFCs were transduced with a tdTomato-expressing lentiviral vector, and the transduction rate reached 85.1% at an MOI of 10 by flow cytometry analysis of transduced and non-transduced cells (Figure 5A). We tested the compatibility of CVECFCs with SIS biomaterial scaffold for future potential tissue engineering. CVECFCs adhered well to the surface of the SIS scaffold, and displayed normal morphology, similar to CB-ECFCs. These results show that CV-ECFCs can be delivered to a defective siteviaa SIS-ECM scaffold or other similar collagen-based scaffolds (Figure 5B).
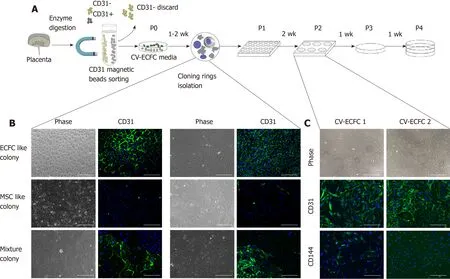
Figure 2 Isolation procedure of endothelial colony-forming cells from placental chorionic villi. A: Depiction of the isolation of chorionic villus endothelial colonyforming cells from chorionic-villus cells by enzymatic dissociation of chorionic villus tissue, followed by CD31 magnetic bead sorting and subsequent manual clonal isolation and expansion; B: Morphology of the colonies formed by CD31+-sorted cells. CD31 was expressed only by colonies that had a cobblestone-like morphology and not by ones that displayed spindle shaped morphology. Two representative colonies are shown for each morphology; C: Immunofluorescence staining of isolated colonies for expression of CD31 and CD144. Scale bar = 100 μm.
DISCUSSION
The placenta is a highly vascularized organ, and the vascularization process begins at an early gestational age[35]. This study confirmed that endothelial cells account for about 20%-40% of total placental cells present during early gestation. We showed that endothelial cells are present in the chorionic villi and likely involved in the formation and development of blood vessels. Proliferative ECFCs are rare in placental cell populations, which makes the isolation of viable ECFCs technically challenging,especially since the number of available cells from clinical chorionic villus sampling(CVS) specimens is limited[40,41]. Flow cytometry sorting is one method that has been used to isolate subpopulations from mixed cells and tissues. However, the viability of flow cytometry-sorted cells is predominantly low, making it unfeasible to obtain large numbers of cells from limited amounts of tissue[41]for anyin uteroautologous fetal therapy. Hence, to isolate a viable and expandable purified population of endothelial progenitor cells (CV-ECFCs), we first utilized CD31 magnetic bead sorting, which resulted in two distinct populations of cells with high proliferative potential. One population had an endothelial progenitor cobble-stone like morphology and coexpressed CD31 and CD34, and are therefore most likely CV-ECFCs. The second population had a mesenchymal spindle shape morphology, and unlike PMSCs that are CD31-negative, these cells co-expressed CD31 and CD90. These cells had a higher proliferative capacity compared to CV-ECFCs, and when co-existing with CV-ECFCs,they outnumber CV-ECFCs. This observation was consistent with the findings of Rappet al[42], where they isolated and cultured ECFCs from term placentas. The recent study showed the presence of bipotent progenitor cells in term placenta, where the CD31Lopopulation differentiated into both endothelial and mesenchymal colonies[40].Hence, CD31/CD90-double positive cells, with the mesenchymal phenotype in our isolation method, could be these bipotential progenitor cells. In addition, according to our previous study, PMSCs could transform into the mesenchymal phenotype and undergo growth arrest when they were co-cultured[20]. Studies have reported that TGF-β can promote the endothelial-to-mesenchymal transition process to make endothelial cells gain a mesenchymal phenotype and induce growth arrest[43,44].Therefore, we added TGF-β inhibitor to the ECGM-MV2 medium to maintain the endothelial cell phenotype. Due to the above reasoning, in order to separate these two cell populations, we picked colonies manually based on their morphology. The expanded cells were further characterized for surface expression of endothelial markers, leading to a pure ECFC cell line that was obtained within 8 wk of culture.
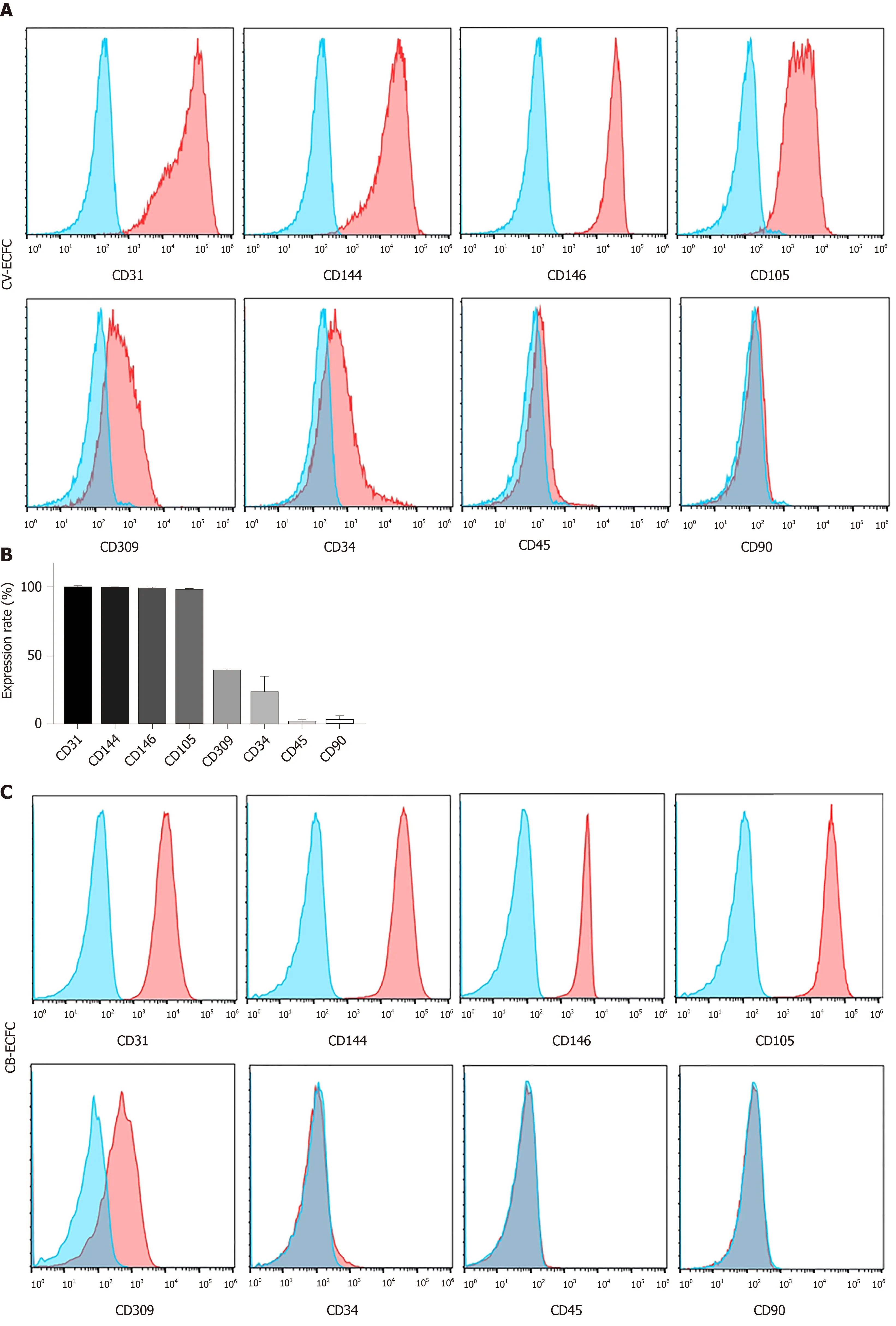

Figure 3 Chorionic villi-derived endothelial colony-forming cells express typical endothelial surface markers, similar to cord blood-derived endothelial colony-forming cells. A: Flow cytometry immunophenotypic analyses demonstrate that chorionic villus endothelial colony-forming cells were positive for the endothelial markers CD31, CD144, CD146, CD105, CD309, low expression of CD34, and were negative for the hematopoietic and MSC markers CD45 and CD90,respectively; B: Quantification of the markers; C and D: Flow cytometry immunophenotypic analysis of the surface marker of cord blood-derived endothelial colonyforming cells. Data are expressed as mean ± SD, n = 3.
One key element of developing well-vascularized, viable regenerative therapeutics is the incorporation of autologous or allogeneic endothelial cells or endothelial progenitor cells derived from various tissue sources to the therapeutic modality or construct. This strategy has been widely used in treating vasculogenesis and/or angiogenesis-related diseases and conditions such as heart failure, acute kidney injury, stroke and wound healing[45-48]. CB-ECFCs are a good source of postnatal treatments, but cannot be used as an autologous cell source for fetal treatments. In this study, we developed a protocol to derive CV-ECFCs from early gestation placentas that could potentially be used as an autologous source of cells for fetal treatment. Our approach allows one to obtain CV-ECFCs from small amounts of tissue (about 100 mg), which is similar to the size of clinical CVS specimens. This method can also be applicable to large-scale expansion and banking of CV-ECFCs from large or whole placental tissue. CV-ECFCs are similar to CB-ECFCs with respect to cell phenotype,in vitrotube-forming capability, transducibility, and compatibility with biomaterial scaffolds. Compared with CB-ECFCs, the CV-ECFCs we isolated express higher levels of the stem cell marker CD34, which likely correlates with a more primitive function and therapeutic capacity. Finally, in combination with the current fetal surgery techniques, these cells hold promise as a novel autologous and/or allogeneic regenerative treatment for congenital anomalies or defects. Furtherin vivofunctional evaluation of these cells is warranted.
In summary, we established a CD31 magnetic sorting-assisted approach to isolate clonal CV-ECFCs from human early gestation placentas that hold promise to be used as an autologous cell source for fetal treatment of congenital anomalies or defects.
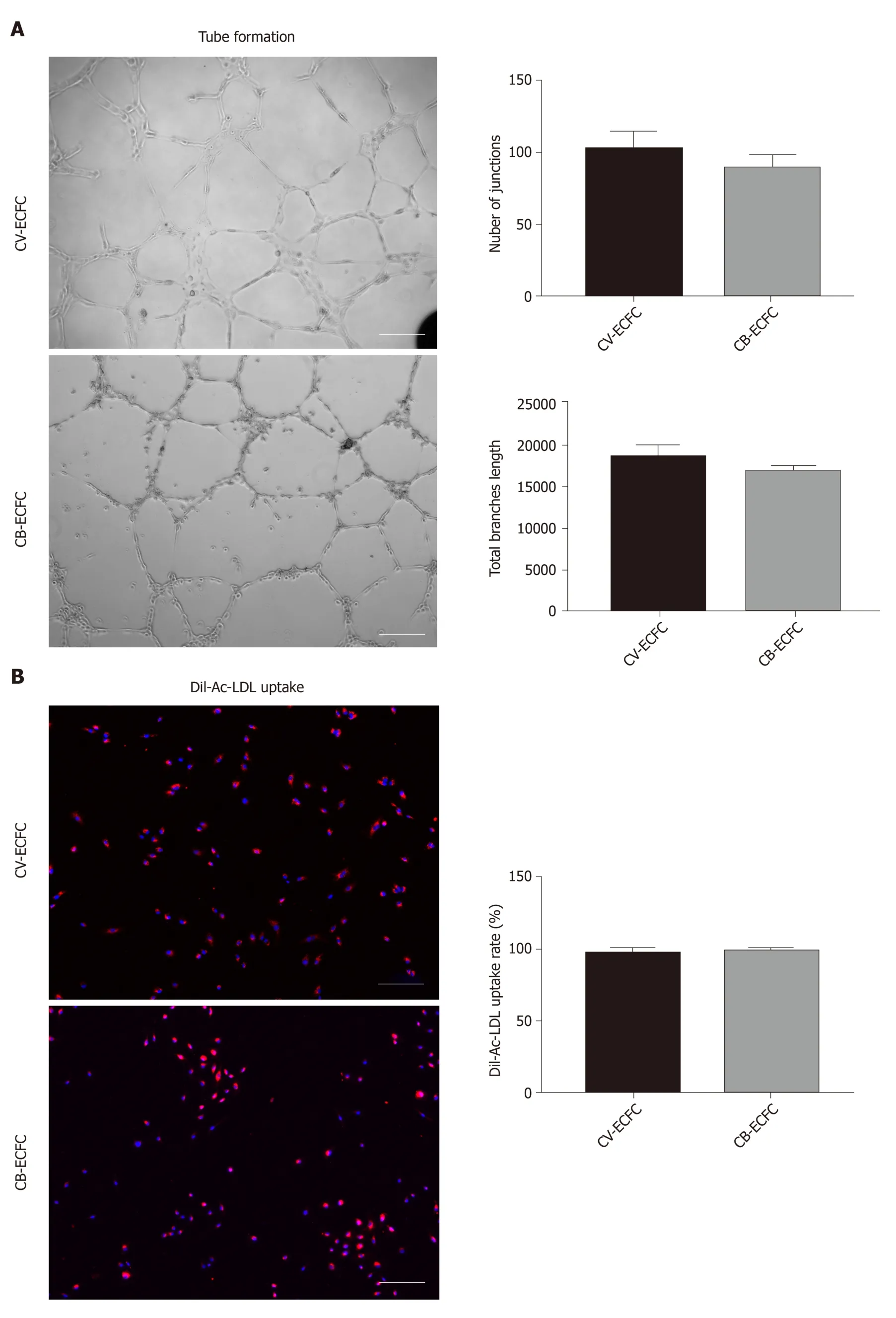
Figure 4 Chorionic villus endothelial colony-forming cells have endothelial functions. A: Representative phase-contrast images of chorionic villus endothelial colony-forming cells (CV-ECFCs), cord blood-derived ECFCs (CB-ECFCs) tube formation assay, and quantification of junction numbers and total branch lengths.There is no difference between CV-ECFCs and CB-ECFCs; B: Representative images for Dil-Ac-LDL uptake by CV-ECFCs and CB-ECFCs. There is no difference in their respective uptake rates. Data are expressed as mean ± SD, n = 3, Scale bar = 100 μm.
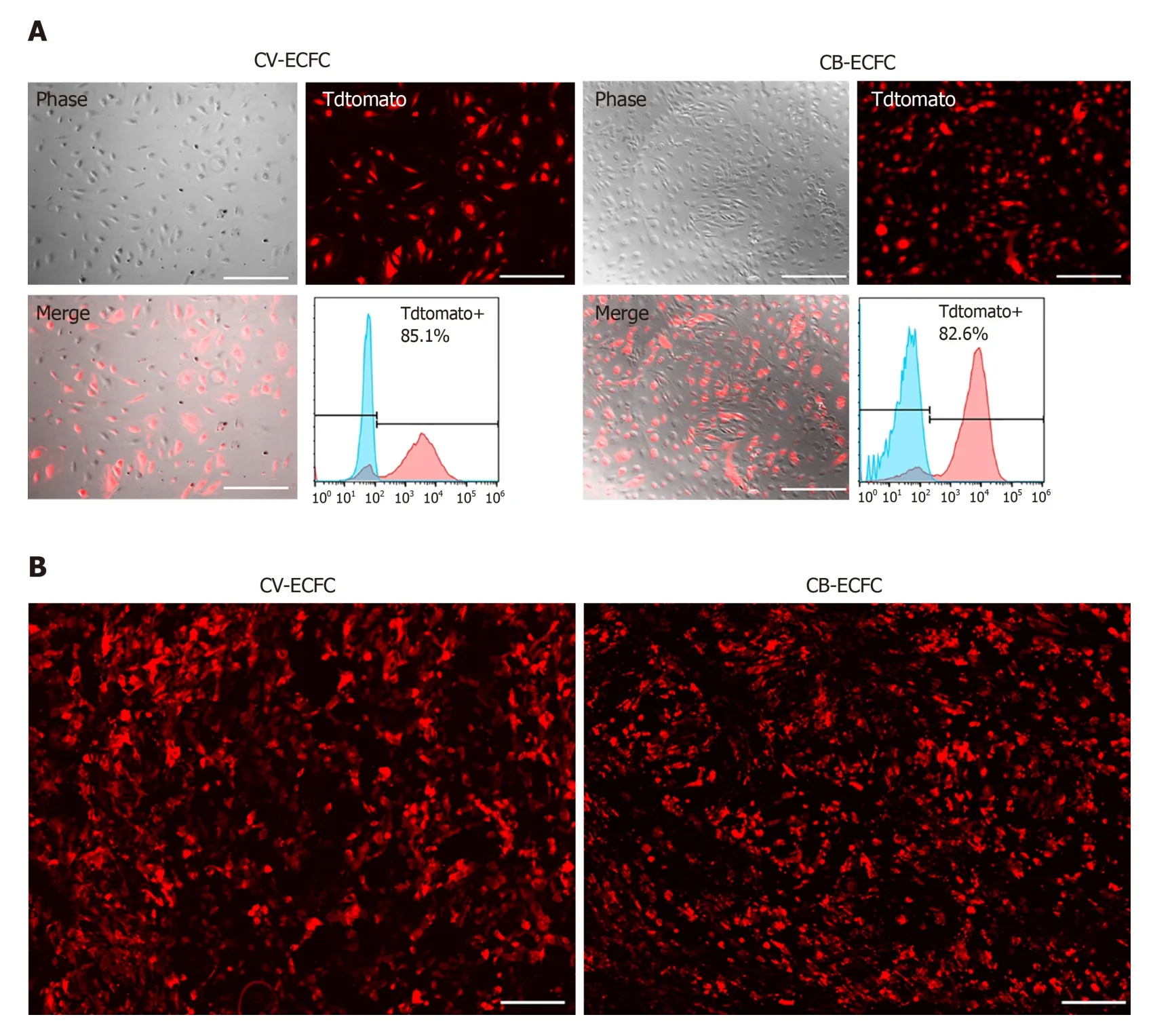
Figure 5 Chorionic villus endothelial colony-forming cells can be transduced by lentiviral vector and are compatible with a small intestinal submucosa scaffold. A: Representative images of chorionic villus endothelial colony-forming cells (CV-ECFCs) and cord blood-derived ECFCs (CB-ECFCs) transduced with a pCCLc-MNDU3-LUC-PGK-Tomato-WPRE lentiviral vector. Flow cytometric analysis showed a transduction rate of 85.1% and 82.8%, respectively; B: tdTomato lentiviral vector-transduced CV-ECFCs (left panel) and CB-ECFCs (right panel) seeded onto a small intestinal submucosa extracellular matrix scaffold, showing adherence. Scale bar = 100 μm.
ARTICLE HIGHLIGHTS
Research background
Fetal medicine and fetal surgery have been substantially developed for the treatment of congenital defects. Stem cell transplantation is an important means of tissue reconstruction and repair of genetic defects. Endothelial colony-forming cells (ECFCs) represent a promising cell candidate for their unique role in facilitating the formation of angiogenesis and vascularization.
Research motivation
ECFCs isolated from the chorionic villus tissue of early gestation placentas can serve as a source of cells for prenatal autologous fetal cell transplantation, as well as provide a basis for studying the development, physiological function, congenital disease, and fetal treatment of the developing fetus and placenta.
Research objectives
The objective of this study is to establish an isolation protocol to obtain ECFCs from the chorionic villus of human early gestation placentas, as well as to investigate the characterization of these cells and their potential applications in gene delivery and tissue engineering.
Research methods
Dissected chorionic villus tissues were enzymatically digested to obtain single cells. Then,magnetic bead sorting, monoclonal culture and colony isolation were performed to obtain chorionic villi-derived ECFCs (CV-ECFCs). Immunohistochemical identification, flow cytometry,Matrigel tube formation assays, LDL uptake assays and lentiviral transduction were carried out to characterize the morphology, phenotype and function of the purified and expanded CVECFCs.
Research results
Using the established isolation protocol, we were able to obtain 1.8 × 107pure CV-ECFCs from a single cell colony culture within 6-8 wks. CV-ECFCs showed typical endothelial phenotypes and functions. CV-ECFCs have demonstrated the ability to be transduced with lentiviruses, and function as carriers for gene therapy. They also possess good biocompatibility with biomaterial delivery vehicles, such as small intestinal submucosa extracellular matrix for potential tissue engineering applications.
Research conclusions
This study shows that ECFCs are present in early gestation placental chorionic villi, and can be isolated and expanded to a significant number in a short period of time. These CV-ECFCs possess typical endothelial cell phenotypes and functions, and hold the potential of being used in gene therapy and tissue engineering applications.
Research perspectives
CV-ECFCs isolated from early gestation placentas provide a new source of ECFCs for the fetal treatment of congenital disorders. Combined with existingin uterotreatment technologies, this cell therapy could be widely applied toward a variety of diseases and conditions. In future research, we will further explore thein vivoapplications of these cells in various animal models.Investigating the phenotype and functions of CV-ECFCs will also facilitate our understanding of the development, cellular composition, and function of the developing placenta and its interaction with the developing fetus.
ACKNOWLEDGEMENTS
The authors thank Cook Biotech Inc. for generously providing us with the ECM material. We acknowledge Alexandra Maria Iavorovschi for the help with manuscript editing.
 World Journal of Stem Cells2020年2期
World Journal of Stem Cells2020年2期
- World Journal of Stem Cells的其它文章
- C-C chemokine receptor type 2-overexpressing exosomes alleviated experimental post-stroke cognitive impairment by enhancing microglia/macrophage M2 polarization
- Comparison between the therapeutic effects of differentiated and undifferentiated Wharton's jelly mesenchymal stem cells in rats with streptozotocin-induced diabetes
- Cartilage and bone tissue engineering using adipose stromal/stem cells spheroids as building blocks
- Mesenchymal stem cells from different sources and their derived exosomes: A pre-clinical perspective