
甘肅慶陽晚中新世板齒犀類的骰骨材料及其形態(tài)學意義
2022-03-03 00:55:22張曉曉孫丹輝
古脊椎動物學報(中英文) 2022年1期
張曉曉 孫丹輝
(1 天津自然博物館 天津 300201)
(2 中國科學院古脊椎動物與古人類研究所,中國科學院脊椎動物演化與人類起源重點實驗室 北京 100044)
(3 中國科學院大學 北京 100049)
1 Introduction
Ringstr?m (1924) erected the elasmothere species,Sinotheriumlagreliion the basis of cranial and mandibular fragments and isolated teeth from Late Miocene deposits in Baode County, Shanxi Province, China.Sinotheriumis the closest genus to the PleistoceneElasmotheriumin terms of dental morphology, so this genus has often been considered to be ancestral to the genusElasmotherium(Prothero et al., 1989; Antoine, 2003).However, early elasmotheres have weak or strong nasal horns, and the transition from a nasal horn to a frontal horn in elasmotheres is not fully understood (Deng et al., 2013a).Deng et al.(2013a) described a skull (IVPP V 18539) found in the Huaigou locality, Linxia Basin, Gansu, which proved thatS.lagreliihas a distinctive frontal structure which shows a morphological intermediate in the nasal-to-frontal horn transition of elasmotheres.Therefore,Sinotheriumhas been regarded as a critical transitional lineage in the evolution of elasmotheres.Specimens ofSinotheriumwere found in Baode, Shanxi (Ringstr?m, 1924); Fugu, Shaanxi (Xue et al., 1995); the Linxia Basin, Gansu (Deng et al., 2013a); Kalmakpai, Kazakhstan (Beliajeva, 1954, 1971; Bayshashov, 1986, 1993); and Chono-Khariakh, Mongolia (Kondrashev, 2000).Recently, a very large rhinoceros cuboid was discovered in a classical paleomammalogy collection, the Licent Collection, which is housed in Tianjin Natural History Museum, Tianjin, China.The specimen was collected from Qingyang, Gansu Province by Emile Licent.
French Jesuit and naturalist Emile Licent collected abundant fossils of theHipparionfauna in 1920 in Qingyang, Gansu Province, China.Most of the mammalian fossils were collected from two localities: Xingjiagou and Zhaojiacha (Licent, 1936).Qiu et al.(1979) indicated that the lithological characteristics of both localities are the same and the two localities have similar stratigraphic positions.From this collection, three subspecies (Adcrocutaeximia variabilis,Ictitherium robustum gaudryi,Ictitherium hipparionum hyaenoides) and four species (Ictitherium wongii,Palinhyaena reperta,P.imbricate,Lycyaena spathulata) of hyenas (Qiu et al., 1979), three species (Hipparioncoelophyes,H.hippidiodus,H.ptychodus) of threetoed horse (Qiu et al., 1987; Sun B et al., 2018), one species (Chilotherium licenti) of rhino (Sun D et al., 2018) and two species (“Gazella”paotehensis, “Gazella”dorcadoides) of gazelle (Li et al., 2018) have been described and published.Of these, the combination ofHipparionandChilotheriumstrongly support a Late Miocene age (Deng, 2006).
Materials and methodsThe study specimen (THP 04553), which is a left cuboid, is housed in the Tianjin Natural History Museum.The specimens for comparison include THP 20304, THP 20306, THP 20307, THP 20309 and THP 07774, which are also housed in Tianjin Natural History Museum, AMNH 129963, AMNH 129964, AMNH M 51855, AMNH M 81805, AMNH M 54456, and AMNH M 81892, which are housed in American Museum of Natural History, and HLMD F 49, which is housed in Hessisches Landesmuseum Darmstadt.
The terminology and measurements follow those of Guérin (1980) and Hünermann (1989).All measurements were performed using calipers (in mm).Measurements in figures and table: 1.maximal anteroposterior diameter; 2.maximal transverse diameter; 3.maximal height; 4.anteroposterior diameter of proximal articular facet; 5.transverse diameter of proximal articular facet; 6.height of anterior part (Fig.1).
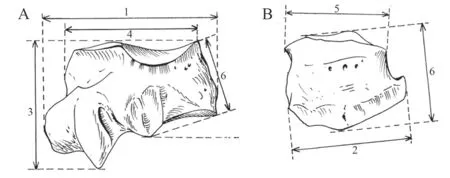
Fig.1 Measurements of rhinocerotid cuboid in lateral (A) and dorsal (B) views Modified from Guérin (1980)
Institutional abbreviationsAMNH, American Museum of Natural History, New York, USA; AMNH M, extant mammalian specimen of AMNH; HLMD, Hessisches Landesmuseum Darmstadt, Darmstadt, Germany; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; THP, Tianjin Natural History Museum, Tianjin, China.
2 Systematic paleontology

DescriptionSpecimen THP 04553 (Fig.2) is a left cuboid, with a trapezoid outline as seen in lateral or mesial view, a remaining damaged surface indicates the presence of a robust posterior apophysis.It is a coarsely parallelepipedal stocky bone.The main part of the bone articulates proximally with the calcaneus and astragalus.These two facets are divided by a very weak central crest.The medial side carries an articular assembly in which five facets can be distinguished: an anterior superior, relatively very small, difficult to distinguish from the proximal joint, corresponding to the navicular; a postero-superior, corresponding to the navicular, which follows the previous one without transition; an upper posteromedian that also corresponds to the navicular; a lower posteromedian, adjoining the previous facet and with which it forms an acute angle; and an antero-inferior semicircular that corresponds to the great ectocuneiform.The distal articular surface is large and concave, and a wide, shallow groove is situated between the surface and the apophysis for the plantar ligament.

Fig.2 Left cuboid of ?Sinotherium, THP 04553 from Qingyang in dorsal (A), plantar (B), proximal (C), distal (D), lateral (E) and medial (F) views
3 Comparison

Fig.3 Ratio diagrams of cuboids of Qingyang specimen and other rhinocerotids The numbers on the x axis are the measurements listed in Fig.1 The y axis is the logarithm (base 10) of the ratios between the measurements of each species and the reference line of zero represents the black rhinoceros, Diceros bicornis
The size of the cuboid from Qingyang is very large, and it is significantly larger than that of any extant rhinocerotid species based on our observations (Fig.3; Table 1).Deng and Zheng (2005) described postcranial material ofElasmotherium caucasicumfrom the Nihewan Basin.We have examined the cuboid specimens described by Deng and Zheng (2005), and found that the Qingyang material is even slightly larger than the average size of the Nihewan specimens.Thus, the Qingyang material belongs to an individual of a huge rhinocerotid taxon, which could be one of the largest rhinocerotids.In addition to the similarity in size, the morphology of the Qingyang specimen is identical to that of the NihewanE.caucasicum, such as the robust bone body and the isosceles trapezoid-shaped outline in lateral view.Although the posterior apophysis is broken, the base of this apophysis is very robust, which is also identical to the morphology of the Nihewan specimens.Thus, the Qingyang specimen can be identified as an elasmothere, or Elasmotheriini.As in the description of Deng and Zheng (2005), the proximal surface is separated by a continuous central crest into two facets for the calcaneus and astragalus.This crest is weak in the Qingyang specimen.Based on observations on specimens of extant taxa, this central crest on the proximal surface is generally strong; thus, the weak crest should be regarded as a primitive character.This is consistent with the fact that the Qingyang specimen was found in the Late MioceneHipparionfauna in the Qingyang area.The Qingyang specimen is most probably attributable toSinotherium, a Late Miocene giant elasmothere genus, which has been found in Baode, Shanxi Province and Linxia Basin, Gansu Province.
Hispanotherium tungurenseis a primitive elasmothere species that lived during the late Middle Miocene (MN7/8) (Cerde?o, 1996).The cuboid ofH.tungurenseis much smaller than those of the Qingyang and Nihewan specimens.In the description by Cerde?o (1996), the posterior apophysis of the cuboid ofH.tungurenseis weakly developed with a slight posterior projection.Indeed, the apophysis of the cuboid ofH.tungurenseis not as strong as that ofElasmotherium caucasicum.The degree of posterior projection of the apophysis ofH.tungurenseis significantly less than that ofE.caucasicum.Although the apophysis is damaged and lost in the Qingyang cuboid, on the basis of the remaining basal part, the angle of posterior projection of the apophysis of the Qingyang specimen was similar as Nihewan ones.The central crest on the proximal surface is weaker than that ofE.caucasicum, similar to that of the Qingyang specimen.This feature is identified as a shared primitive character of early elasmotheres, such as central crest on the proximal surface is relatively weak.
The subfamily Aceratheriinae is another important extinct Miocene taxon.Two taxa, Chilotheriini and Aceratheriini, were widespread in Eurasia during the Late Miocene.Aceratheriini consists of two main genera,AcerorhinusandAceratherium.Most species ofAcerorhinusoccur in China (Deng, 2000; Deng and Chen, 2015).Three valid species ofAceratheriumhave been confirmed (Deng et al., 2013b).We have had the opportunity to observe cuboid material of representatives of both tribes; specifically, of the speciesChilotheriumwimani(from Linxia Basin, Gansu, China) (Deng, 2001) andAceratheriumincisivum(from H?wenegg, Germany) (Hünermann, 1989).The cuboid of Aceratheriinae possesses a unique character that is distinct from that of any other rhinocerotid taxon.The main part of the cuboid of this subfamily has a diamond or parallelogram-shaped outline in lateral view, different from the trapezoid or rectangle-shaped outlines of other taxa.The posterior apophysis of the cuboid of Aceratheriinae is very strong, and forms a considerable proportion of the whole bone body.The basal part of the apophysis is highly inflated.The proximal articular facets for the calcaneus and astragalus and the basal part of the apophysis will all be visible in proximal view on a cuboid of Aceratheriinae.In other taxa, however, only the articular facets can be seen in proximal view.

Table 1 Comparison of cuboids of elasmotheres in China (mm)
Rhinocerotini is regarded as the most advanced rhinocerotid taxon, and is usually referred to as the “true rhinos”.This tribe contains all the extant species and some extinct ones, such asCoelodonta, an iconic woolly rhino of the Ice Age.However, in comparison with other tribes, the morphology of the cuboid of Rhinocerotini is variable.The sizes of the cuboids of all rhinocerotids are smaller than those of the Qingyang and Nihewan specimens.The cuboid size of the smallest extant species,Dicerorhinus sumatrensis, is similar to that ofAceratheriumincisivum.The relative sizes of the apophysis of extant species are generally larger than those of elasmotheres.InRhinoceros unicornisandDiceros bicornis, the main part of the cuboid and apophysis are divided into two distinct parts by a deep groove.The apophysis is even situated completely posteriorly on the cuboid ofD.bicornis.However, there is almost no clear morphological similarity of the cuboid shared by Rhinocerotini.The phylogenetic relationships of the Rhinocerotidae are still controversial.Deng and Chen (2015) proposed that Elasmotheriini and Rhinocerotini are sister groups.Within the Rhinocerotini, Deng et al.(2011) suggested thatCeratotherium simumandD.bicornisoriginated from one clade, whereasDicerorhinus sumatrensis,RhinocerosandCoelodontawere derived from another.Lu (2014) argued thatDicerorhinus sumatrensisandRhinocerosform a monophyletic group andC.simumandD.bicornisform a clade.TheDicerorhinus sumatrensis-Rhinocerosclade and theCeratotherium simum-Diceros bicornisclade are the sister groups of another clade consisting of two elasmothere species,ElasmotheriumsibiricumandNingxiatherium longirhinus.Antoine (2002) argued that all the extant species andCoelodonta antiquitatisform a monophyletic group.The variable morphological features of the cuboids of Rhinocerotini suggest complicated phylogenetic relationships.
Teleoceratini is a distinctive taxon of the Rhinocerotidae.They are hippo-like animals that lived close to rivers (Prothero, 2005).Their cuboids are also highly specialized with a very low dorsoventral height with an inflated apophysis with a strong posterior projection.This morphological pattern is completely distinct from those of other taxa of the Rhinocerotidae (Guérin, 1980).

Fig.4 Reconstructed superficial muscle-ligament system of the hind limb of rhinocerotoid, based on Hünermann (1989), Qiu and Wang (2007)
4 Discussion
Deng et al.(2012) proposed that some morphological features of equid postcranials may have allowed the animal to stand more vertically.This is an adaption for an open habitat in a cold and arid environment; indeed this phenomenon can be observed within the majority of ungulates.Similar features with functional significance, especially for standing and running, are also present in rhinos, for example, as documented in the work of Qiu and Wang (2007) on the muscle-ligament system and functional analysis of paraceratheres.In their reconstruction of the superficial muscles ofJuxia sharamurenensis, they suggested that the flexor digitalis pedis profundus muscle, flexor digitalis pedis superficialis muscle and peroneus muscle are attached to the posterior apophysis of the cuboid.The ligaments need to pull the foot effectively in the upward direction, so the main axis of the apophysis must be nearly vertical to the horizontal direction (Fig.4).The smaller the angle between the frontal plate of the main part and the main axis of the apophysis on the cuboid, the more vertically the foot can stand (Fig.5).In the present research, we chose cuboids from three elasmothere species (Hispanotherium tungurense,Elasmotherium caucasicumand Qingyang specimen),Coelodontaantiquitatisand four extant species (Ceratotherium simum,Diceros bicornis,Rhinoceros unicornisandDicerorhinus sumatrensis).Below, the degree between the frontal plate of the main part and the main axis of the apophysis on the cuboid for several rhino species and the habitat preferences of those species (information of all the extant species after Wilson and Mittermeier (2011)) are compared (Fig.6).
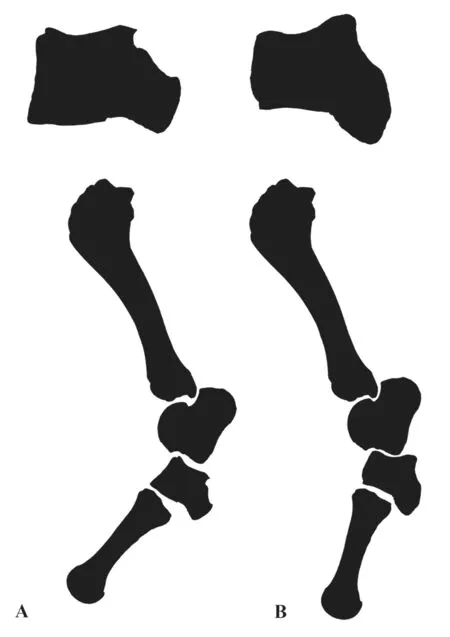
Fig.5 Diagram of the hind limb skeleton showing the position of the cuboid in relation to the other bonesA.more flexed limb posture; B.less flexed limb posture.Modified from Hünermann (1989)
Both the northern and the southern subspecies ofCeratotherium simum(white rhinoceros) are grazers, and are restricted to grassland and savanna habitats.They prefer to live in areas where rainfall levels are intermediate (around 700-800 mm).Diceros bicornis(black rhinoceros) can be found in many habitats in Africa.This species occurs primarily in mesic wooded savanna, but can be found in wetter montane forests in Kenya, wetter dune forests in eastern South Africa, and semi-deserts in Kunnene, Namibia.Rhinoceros unicornis(greater one-horned rhinoceros) flourishes in protected areas along the main river systems in the lowlands of the Himalayas.This species prefers to chooseShorea robustaforest as refugia during monsoon inundations.They are also rarely found more than 2 km from water.Dicerorhinus sumatrensis(Sumatran rhinoceros) have been usually found in a range of environments from lowland semi-evergreen forests to highelevation cloud forests.The habitat information of all the extant species above is consistent with the morphologies of their cuboids.The angle between the frontal plate of the main part and the main axis of the apophysis on the cuboid is smallest inCeratotherium simum, so its ability to stand vertically is very strong, representing an adaptation to its relatively arid habitat.The other three species all live in moist habitats with abundant water systems or wood.
Deng et al.(2011) suggested that all of the woolly rhinos lived in cold environments, for example, in northern Eurasia, especially Siberia.The ancestral woolly rhino,Coelodontathibetana, descended from the Tibetan Plateau to northern latitudes and turned to grazing in the Late Pleistocene as global climate cooled and cold habitats expanded, eventually becoming one of the most successful Ice Age mega herbivores.Hence, the habitat ofC.antiquitatisshould have had the highest degree of openness and aridity.This finding perfectly fits with the degree crossed by the frontal plate of the main part and the main axis of the apophysis on the cuboid ofC.antiquitatis.This angle is smallest inC.antiquitatisamong all of the species considered herein.
The degrees crossed by the frontal plate of the main part and main axis of the apophysis on the cuboid are similar amongCeratotherium simum,AceratheriumincisivumandHispanotherium tungurense, suggesting that the habitats of these two extinct mediumsized rhinos were similar to that ofCeratotherium simum.It has been proposed that global warming led to the Middle Miocene Climatic Optimum (MMCO) (Zachos et al., 2001; Deng, 2016).Hispanotherium tungurensewas found in the Tunggur fauna, which lived during the late Middle Miocene (MN7/8) in Nei Mongol, China.The environment was significantly drier than present conditions between 13.2 and 11.5 Ma (Serravallian), with typically less than 50% precipitation compared to recent measurements, representing the most intense and prolonged arid period in Europe since the Middle Miocene climatic optimum (B?hme, 2003).Aceratheriumincisivumwas found in the H?wenegg fauna (MN9-10), which lived in Germany during the early Late Miocene.The elapsed time from the end of the MMCO to these faunas was long, so these two species probably lived in relatively open habitats.
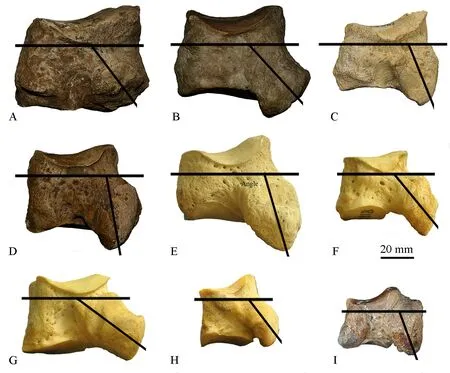
Fig.6 Morphological comparison of cuboids of the Qingyang specimen and other rhinocerotids, all specimens are left and showed in lateral viewA.THP 04553, ?Sinotherium in Qingyang; B.THP 20309, Elasmotherium caucasicum in Nihewan; C.AMNH 129963, Hispanotherium tungurense in Tunggur; D.THP 07774, Coelodonta antiquitatis in Sjaraosso-gol; E.AMNH M 51855, Ceratotherium simum; F.AMNH M 81805, Diceros bicornis; G.AMNH M 54456, Rhinoceros unicornis; H.AMNH M 81892, Dicerorhinus sumatrensis; I.HLMD F 49, Aceratherium incisivum in H?wenegg
The most interesting question comes from the two giant elasmothere species.Deng and Zheng (2005) argued thatElasmotherium caucasicumhas hypsodont cheek teeth with thick cement and complicated folds as well as slender limbs, which indicates that this species is a cursorial runner and grazer.Deng et al.(2013a) argued thatSinotheriumhas a low positioned and dolichocephalic head in addition to hypsodont cheek teeth with thick cement and complicated folds, which indicates that this species is a typical grazer.However, the degrees between the frontal plate of the main part and the main axis of the apophysis on the cuboid of the Qingyang and Nihewan specimens are relatively large, even larger than that ofCeratotherium simum, and are similar with those of extant species living in moist habitats.Sinotheriumhas been found in the Huaigou locality, Guanghe County, Gansu.The Huaigou fauna is a typical Late MioceneHipparionfauna, but contains some species that lived in a forest environment, such as chalicothere (Ancylotheriumsp.), deer (Dicrocerussp.) and giraffe (Palaeotragus microdon).The fauna component is the indicator for the presence of forest, which is consistent with the hypothesis that the huge animalSinotheriumlived in a forest habitat like the extant huge animalsElephas maximus(Asian elephant) andLoxodonta cyclotis(African forest elephant) (Wilson and Mittermeier, 2011).Forest representatives were also part of the Nihewan fauna, such asMuntiacus bohlini,Eucladoceros boulei,Cervus elegans,Cervus(Rusa) sp.andElaphurus bifurcates.There are also abundant giraffe species specimens in the Xingjiagou and Zhaojiacha localities in Qingyang, Gansu.So it is not impossible that giant elasmotheres lived in a forest or wooded habitat.
The diversity of elasmotheres in the Linxia Basin is greater than in any other locality.The fauna of the basin contains one primitive species (Hispanotherium matritense(Deng, 2003), and four relatively derived species (Parelasmotherium linxiaense(Deng, 2007);Parelasmotherium simplum(Qiu and Xie, 1998);Ningxiatherium euryhinus(Deng, 2008);Iranotherium morgani(Deng, 2005).The Linxia Basin appears to have been an evolutionary center for elasmotheres.The discovery of cuboid material ofSinotheriumin Qingyang makes the elasmothere dispersal route clearer.It is hypothesized thatSinotheriumoriginated in the Linxia Basin and then dispersed in two directions: one route extended east, via Qingyang, to Baode, Shanxi Province, and Fugu, Shaanxi Province.The other route extended northwest via Mongolia to Kazakhstan.The main distribution area of relatively primitive species ofElasmotheriumis northeast of the Black Sea (present-day Russia and central Asia) (Fig.7).The most advanced species,Elasmotheriumsibiricum, occurs widely in the middle and northern parts of Eurasia.The dispersal ofSinotheriumto northern Asia probably suggests that elasmotheres gradually tended to evolve to become more cold-adapted since the Late Miocene.The Mid-Pleistocene transition (MPT) was a very important climate event during the Quaternary and was initiated by a shift towards much larger Northern Hemisphere ice shields at around 0.9 Ma (Mudelsee and Schulz, 1997; Raymo et al., 1997; Schmieder, 2000).Elasmotheriumsibiricumseemed to adapt well to the cold climate and became widespread in the early stage of the MPT, becoming a common element of the Eurasian Quaternary mammal faunas.

Fig.7 Paleozoogeography of giant elasmotheres Modified from Shvyreva (2015)
AcknowledgementsWe thank Deng Tao and Sun Boyang (IVPP) for their helpful suggestions and for improving the manuscript.Judy Galkin and Eleanor Hoeger (AMNH) respectively provided accesses to observations on the specimens of both fossil and extant rhinos.Oliver Sandrock (HLMD) provided accesses to observations on the H?wenegg collection.Shi Qinqin (IVPP) provided very important literature for references.We appreciate all of their contributions.This work was supported by the Chinese Academy of Sciences (Grant Nos.XDB26030302, XDA20070203).
猜你喜歡
青少年科技博覽(中學版)(2023年1期)2023-03-17 00:44:24
科學大眾(2022年23期)2023-01-30 07:03:18
河北地質(zhì)大學學報(2021年5期)2021-11-08 06:27:24
考試與評價·七年級版(2021年4期)2021-08-14 21:55:33
甘肅社會科學(2021年3期)2021-06-02 08:56:26
學生天地(2020年19期)2020-06-01 02:11:32
今日農(nóng)業(yè)(2019年14期)2019-01-04 08:57:40
中國繼續(xù)醫(yī)學教育(2015年2期)2016-01-06 01:36:16
大眾考古(2015年5期)2015-06-26 08:21:36
振動、測試與診斷(2014年6期)2014-03-01 01:14:50
