
Seasonal distribution of macrobenthos and its relationship with environmental factors in Yellow Sea and East China Sea*
2018-07-11 01:58SHOULu壽鹿LIAOYibo廖一波TANGYanbin湯雁濱CHENJianfang陳建芳JIANGZhibing江志兵GAOAigen高愛根CHENQuanzhen陳全震
SHOU Lu (壽鹿) LIAO Yibo (廖一波) TANG Yanbin (湯雁濱) CHEN Jianfang (陳建芳)JIANG Zhibing (江志兵) GAO Aigen (高愛根) CHEN Quanzhen (陳全震)
1Key Laboratory of Marine Ecosystem and Biogeochemistry,Second Institute of Oceanography,State Oceanic Administration,Hangzhou 310012,China
2Key Laboratory of Applied Marine Biotechnology,Ministry of Education,Marine College of Ningbo University,Ningbo 315211,China
AbstractMacrobenthos samples were collected from the Yellow and East China Seas in four seasons during 2011 to 2012. The seasonal distribution of macrobenthos and its relationship with environmental factors were analyzed. A total of 562 macrobenthic species were identified, with polychaetes and mollusks accounting for 67% of the total number of species. A similarity percentage (SIMPER) analysis showed that the dominant species were bivalve mollusks in the Yellow Sea and small-sized polychaetes in the East China Sea. A two-factor analysis of variance showed significant seasonal variations in species number,density and diversity index, and significant regional differences of biomass and density. Two-factor community similarity analysis also showed significant seasonal and regional differences in macrobenthic communities. Canonical correspondence analysis indicated that the main environmental factors affecting the macrobenthic communities were water depth, temperature, dissolved oxygen, and inorganic nitrogen. The results demonstrate significant regional differences and seasonal variations in macrobenthos in the two seas.Sediment properties and water mass characteristics are speculated to be the causes of regional differences.
Keyword:Yellow Sea; East China Sea; macrobenthos; environmental correlation analysis
1 INTRODUCTION
Macrobenthos are consumers and transporters in material circulation and energy flow within marine ecosystems, and are important in both pelagic-benthic coupling and biogeochemical cycles (Conlan et al.,2008). The benthos can alter the chemical composition of sediments and affect their stability by bioturbation(Norkko et al., 2001), consume oxygen and release particulate matter and nutrients to the water column through feeding, burrow construction and other activities (Kamp and Witte, 2005), obtain nutrients by consuming plankton (Snelgrove et al., 2001), and be preyed on by demersal fi sh (Crawford et al., 2007).
As one of the world’s most important marginal seas, the Yellow and the East China Seas have attracted widespread attention from scientists (Li, 2003; Liu and Li, 2003; Lin et al., 2004; Peng et al., 2014).Owing to high costs, marine ecological surveys over the past few decades have been limited on both spatial and time scales. For example, most studies have focused on macrobenthos in estuaries (Li et al., 2007a,2016a, b; Liu et al., 2008, 2014), nearshore (Hu et al.,2000, 2006) or bays (Li et al., 2001, 2005a; Liao et al.,2011; Wang et al., 2011; You et al., 2011), with only one or two cruises (Liu and Li, 2003; Lin et al., 2004;Peng et al., 2014) in the Yellow Sea, East China Sea or adjacent waters. There are clear spatiotemporal variations in the environmental characteristics of the Yellow and East China Sea regions, which may lead to incomplete understanding of changes in macrobenthos distribution and corresponding inf l uences.
With the opportunity offered by the National Program on Key Basic Research and Development program entitled “Evolution, mechanism and the prediction of the future trends of the marine ecological environment in the continental shelf of China,” we conducted a four-season survey and sampled macrobenthos of the Yellow and East China Seas during 2011 to 2012. Our study was aimed at illustrating temporal and spatial distributions of macrobenthic communities in the studied areas, to understand the relationship between benthic communities and environmental factors and provide a reference for understanding the evolution of marine ecosystems and environmental effects.
2 MATERIAL AND METHOD
2.1 Study area
The East China and Yellow Seas adjoin at midlatitudes, forming a western boundary of the Pacific Ocean. Because wide continental shelves and large amounts of terrigenous material received from the Changjiang (Yangtze) River, the East China and Yellow Seas have a highly representative ecosystem,and are thus becoming some of the world’s most important marginal seas (Zhai et al., 2014). Rapid development and previous exploitation have changed the seas into the most vulnerable ecosystem in the world. The ecological security of these areas is strongly related to the sustainable development of society and the economy. In recent years, because of rapid economic development, the marine ecology,resources, environment, and other aspects of this area have been damaged to varying degrees by humans,and are under enormous pressure (Anderson et al.,2002; Li et al., 2007b; Jiang et al., 2014). The southern Yellow Sea is mainly inf l uenced by the Yellow Sea Coastal Current, Yellow Sea mixed water, Yellow Sea Cold Water Mass (YSCM), and river plumes from the Han-gang and Huaihe rivers (Peng et al., 2014). The East China Sea, on the other hand, is mainly inf l uenced by Changjiang Diluted Water, Taiwan Warm Current,the Kuroshio, coastal current, YSCM, shelf mixed water and upwelling in the Changjiang River Estuary,and Zhejiang coastal waters (Su and Yuan, 2005).
2.2 Sampling design
Samples were taken during a cruise in the Yellow and East China Seas on R/VDong Fang Hong No. 2during March 17-April 7, 2011 (Spring), July 6-July 24, 2011 (summer), October 17-November 5, 2011(autumn), and December 20, 2011-January 11, 2012(winter). The sampling sites covered most parts of the two seas, with slight adjustments in different seasons(Fig.1). The survey and monitoring were performed according to “The Specification for Oceanographic Survey” (GB/T 12763-2007) and “The Specification for Marine Monitoring” (GB17378-2007). At each sampling site, a Day grab sampler (0.1 m2) was used to collect sediments twice per site, followed by rinsing with a 0.5-mm mesh sieve. Samples were fi xed with 5%-7% neutral buffered formaldehyde in situ, and sample analysis (e.g., species identifi cation, counting and weighing) were performed in the lab. Bottom water temperature, salinity and depth were recorded by a SeaBird 911 CTD (Sea-Bird Electronics,Bellevue, WA, USA). Simultaneously, we measured pH and the concentrations of chlorophylla, dissolved oxygen (DO), phosphate (PO4-P), dissolved inorganic nitrogen (DIN, including nitrates, nitrites and ammonium salts), and silicate (SiO3ˉ) of the bottom water.
2.3 Data processing
A two-factor analysis of variance (ANOVA) was performed using STATISTICA 6.0 on parameters for macrobenthic communities in the Yellow and East China Seas, with season and region as factors. Before statistical analysis, normality and homogeneity of data variance were examined using the Kolmogorov-Smirnov and Bartlett tests, respectively, and the signifi cance level was set toα=0.05 (Yi, 2002).PRIMER 6.0 software was used to calculate the species number (S), Shannon-Wiener diversity index(H'), and Pielou’s evenness index (J) of the macrobenthic communities. Macrobenthic communities in various seasons and regions were compared using two-factor analysis of similarities(ANOSIM), and multi-dimensional scaling (MDS)were used to investigate differences in community structure within PRIMER 6.0. At the same time, the average contribution rate of each macrobenthic species to similarity in the different regions was analyzed by the similarity percentages (SIMPER) method, and the five species with the highest cumulative contribution rates in each region and season were defi ned as the main characteristic species (Clarke and Gorley, 2006).Canonical correspondence analysis (CCA) was done using Canoco for Windows 4.5 to analyze the relationship between dominant macrobenthic species and environmental parameters, and the signifi cances of all ordination axes of CCA were examined using a Monte Carlo test (Ter Braak and ?milauer, 2002).
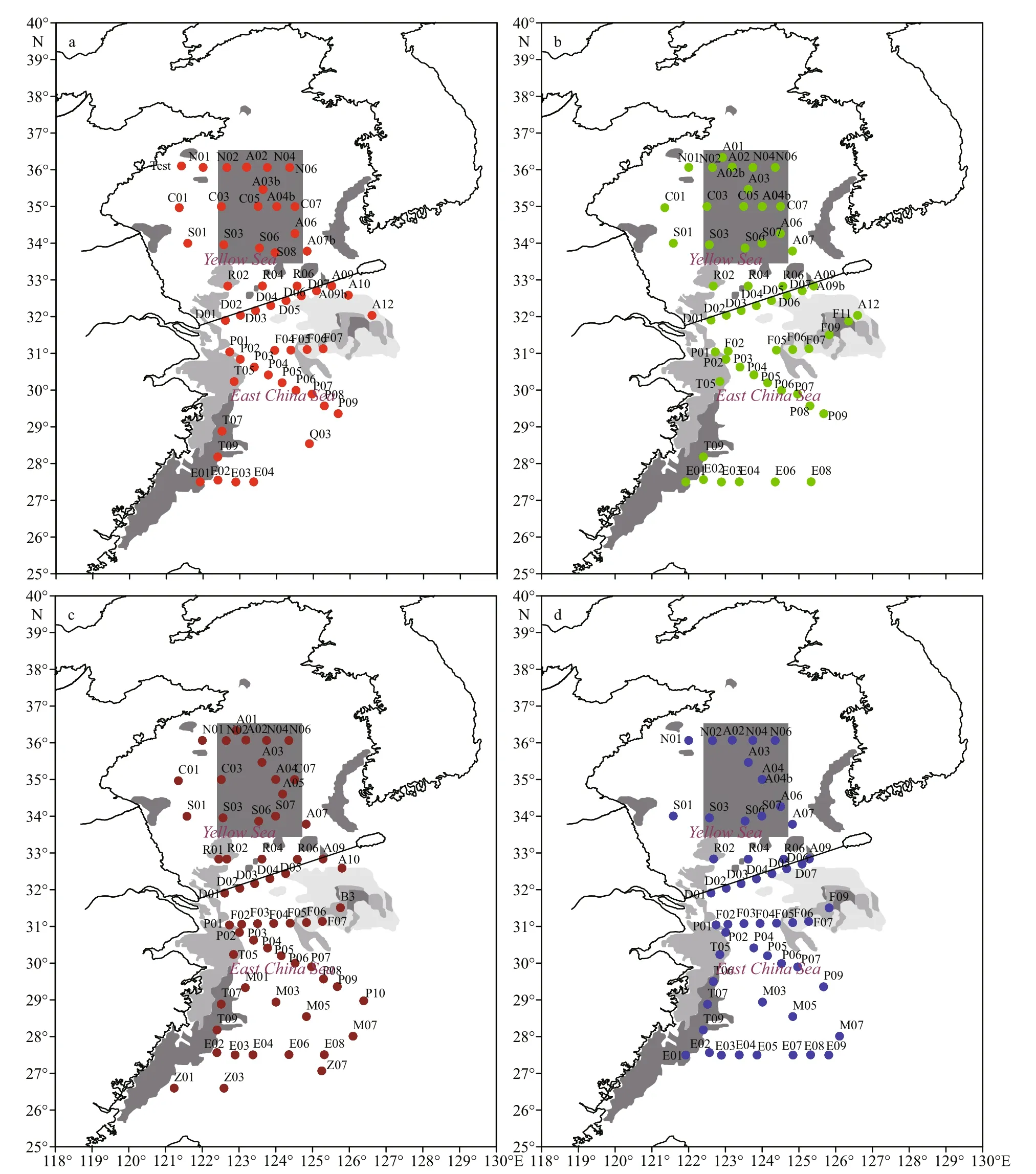
Fig.1 Sampling sites of macrobenthos in Yellow and East China Seasa. spring; b. summer; c. autumn; d. winter.
3 RESULT
3.1 Composition of macrobenthic species and dominant species
3.1.1Species composition and distribution
Among the macrobenthos collected from theYellow and East China seas, 562 species were identified, including 224 of polychaetes and 151 of mollusks, accounting for ~67% of all macrobenthic species, thus constituting the dominant species in the study area. The macrobenthic species compositions in various regions are shown in Fig.2. In the southern Yellow Sea, there were 331 macrobenthic species,including 160 of polychaetes and 72 of mollusks(accounting for ~70% of the total number of all species). In the East China Sea, there were 453 macrobenthic species, including 194 of polychaetes and 114 of mollusks (accounting for ~68% of the total number of all species).
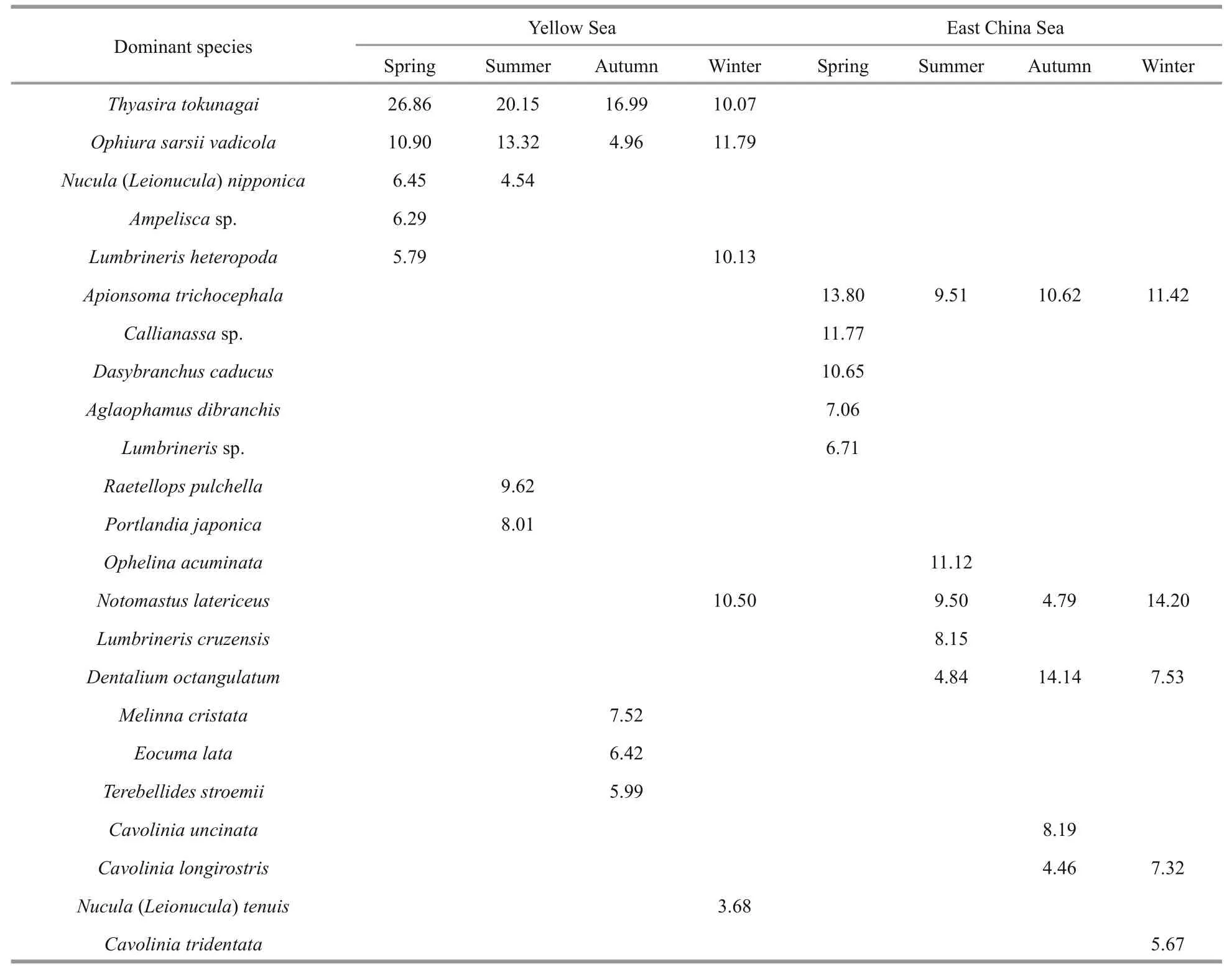
Table 1 Dominant species of macrobenthos in Yellow and East China Seas and their percentage (%) contribution to the community
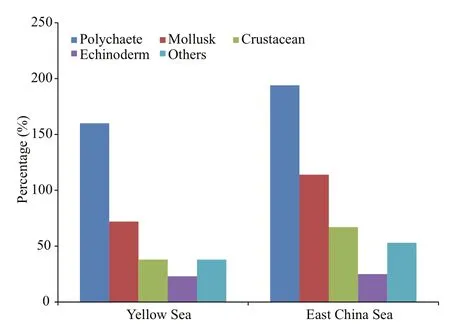
Fig.2 Species composition of macrobenthos in Yellow and East China Seas
3.1.2Dominant species
The result from SIMPER analysis showed that the dominant species in the Yellow and East China seas were significantly different, and the community dissimilarity indexes for four seasons were all >90%(spring: 96.67%; summer: 94.74%; autumn: 93.56%;winter: 92.05%). In all seasons, the dominant species in the Yellow Sea had larger sizes (e.g., bivalves),while those in the East China Sea were smaller (e.g.,polychaetes) (Table 1).

Table 2 Seasonal and regional variation of macrobenthic community parameters in Yellow and East China Seas

Table 3 Eigenvalues for CCA axes and species-environment correlations
3.2 Parameters for macrobenthic communities
The parameters for macrobenthic communities in the Yellow and East China Seas are shown in Table 2.The result from two-factor ANOVA showed that the species number, density and diversity index all had significant seasonal variations. In the Yellow Sea, the species number and diversity index in fall and winter were significantly larger than those in spring and summer, respectively, and the density in fall was significantly higher than those in other seasons. In the East China Sea, the species number, density and diversity index in summer and fall were significantly higher than those in winter and spring, respectively.significant regional differences were evidenced for biomass and density. Except for summer, densities in other seasons and biomass in all seasons in the Yellow Sea were significantly higher than those in the East China Sea.
The two-factor ANOSIM was performed on the macrobenthic communities, with region (the Yellow Sea and the East China Sea) and season (four seasons)as factors. The macrobenthic communities showed significant seasonal (global testR=0.134,P=0.001)and regional (global testR=0.231,P=0.001)differences, as in MDS analysis (Fig.3).
3.3 Relationship between macrobenthos and environmental factors
The dominant species and environmental factors were selected for CCA. The Monte Carlo signifi cance test revealed that the fi rst and all ordination axes in the CCA had significant differences (1staxis:F=12.174,P=0.016; all axes:F=3.921,P=0.002). As seen from the CCA ordination diagram (Fig.4), water depth, temperature, DO and DIN had larger impacts on macrobenthos among the various environmental factors. Among them, temperature was negatively related to the fi rst axis, with a correlation coeffcient of -0.897. DO was positively correlated with the fi rst axis, with a correlation coeffcient of 0.301 4. Water depth was positively correlated with the second axis,with a correlation coeffcient of 0.595 9, and DIN was negatively correlated with the second axis, with a correlation coeffcient of -0.380 4. Table 3 shows characteristic values of each ordination axis and the correlation coeffcients between the macrobenthic functional groups and environmental factors.
4 DISCUSSION
4.1 Differences between macrobenthic communities in Yellow and East China Seas and spatiotemporal variations
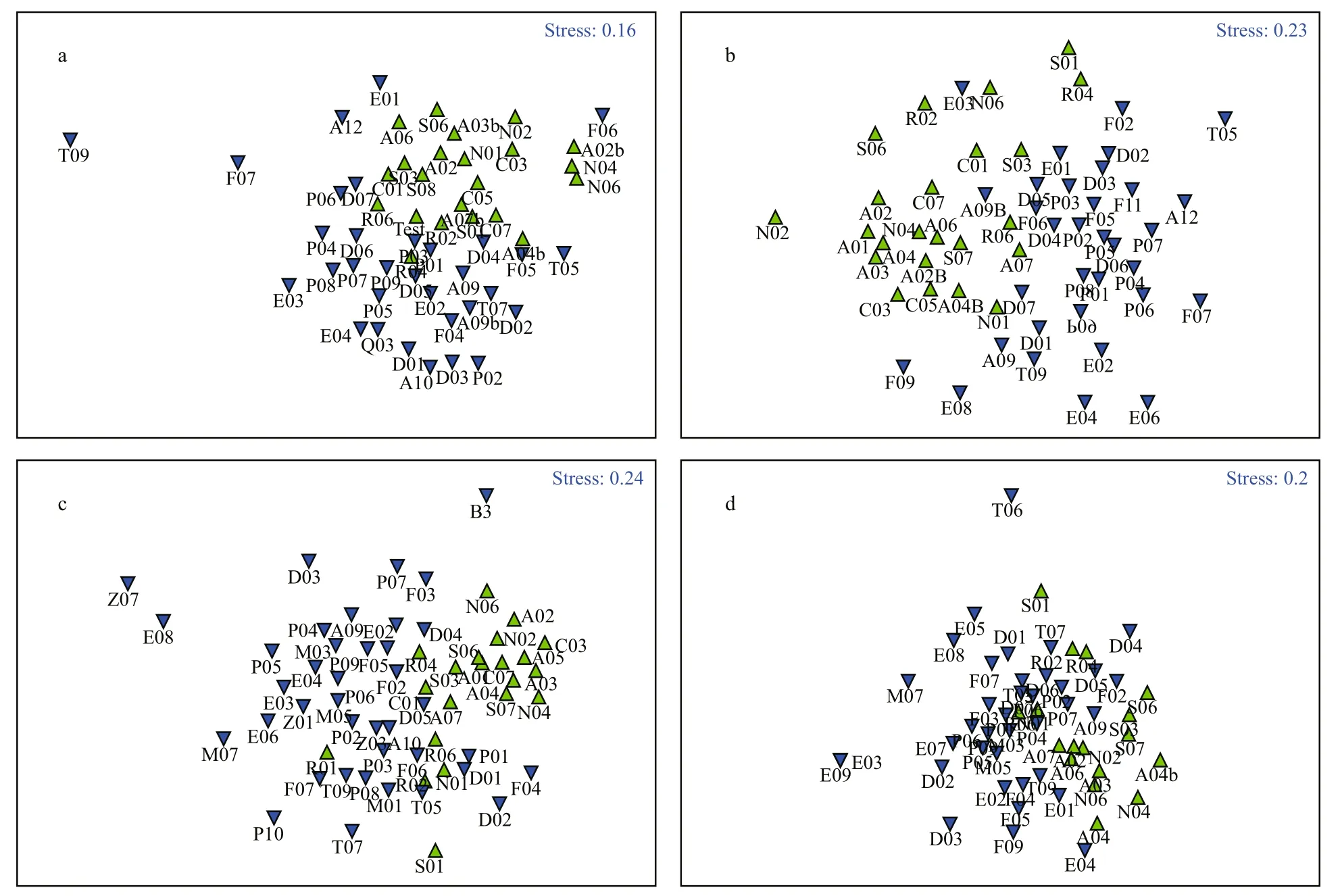
Fig.3 MDS analysis of macrobenthos in Yellow and East China Seasa. spring; b. summer; c. autumn; d. winter; ▲: Yellow Sea; ▼: East China Sea.
Two-factor community ANOSIM results indicated significant differences in macrobenthic community structure across the two seas (global testR=0.231,P=0.001). In terms of species composition, 562 macrobenthic species were identified from the two seas, and the number of such species in the East China Sea (453) was significantly larger than that of the Yellow Sea (331). The dominant macrobenthic species in both seas were polychaetes and mollusks,whose proportions were very similar in the two seas.Liu and Li (2002, 2003) reported that during spring/fall in the East China Sea and southern Yellow Sea,there were a total of 392 and 272 macrobenthic species in 2002 and 2003, respectively, slightly smaller numbers than in the present research because of differences in survey sites and season. Furthermore,except for a box corer (0.1 m2), an Agassiz trawl was used for qualitative sampling of benthic fauna in Liu and Li (2002, 2003), and the proportions of crustaceans (East China Sea: 30.1%; Yellow Sea:30.5%) were much larger than those in our survey(East China Sea: 14.8%; Yellow Sea: 11.5%). In Jiang et al. (1985a), 1287 macrobenthic species were found in the continental shelf of the East China Sea and adjacent areas from October to November 1977 and September to October 1978. Among them, 770 species were identified, much larger than numbers from other survey data over the years. In Li (2003),there were 414 and 855 macrobenthic species in the Yellow and East China seas, respectively. Peng et al.(2014) reported 306 macrobenthic species in the Yellow and East China seas during spring and summer 2011, and the dominant species were polychaetes, crustaceans and mollusks. Different times, frequencies and ranges of surveys, plus use of the Agassiz trawl to sample benthic fauna qualitatively(or not) caused substantial differences between the surveys of macrobenthic species in the two seas.More macrobenthic species were found in surveys with larger area and frequency, and larger proportions of crustaceans were found with use of the Agassiz trawl.
In all seasons, the average density and biomass of macrobenthos in the Yellow Sea was 341.7 ind./m2and 51.31 g/m2, respectively, and these were significantly less in the East China Sea (253.5 ind./m2and 20.02 g/m2, respectively). There was a notable difference in macrobenthos number in the two seas during different survey periods (Table 4). In the Yellow Sea, the average biomass and density of macrobenthos in our survey were greater than historical data of spring and fall. That is, average biomass was 21.51 g/m2in 1959, 43.18 g/m2in 1998-2000, 30.35 g/m2in 2000-2001, and 44.17 g/m2in the present survey. Average density was 261 ind./m2in 1998-2000, 156.5 ind./m2in 2000-2001, and394.7 ind./m2in our survey. In the East China Sea during spring and fall, the average biomass of macrobenthos was 33.46 g/m2in 1959, 30.17 g/m2in 1998-2000, and 8 g/m2in 2000-2001. Average density was 398.5 ind./m2in 1998-2000 and 117.3 ind./m22000-2001. In the present survey, the average biomass of spring and fall was 19.61 g/m2,smaller than the value of 1959 and 1998-2000, but larger than that of 2000-2001. Average density was 248.5 ind./m2, smaller than the value of 1998-2000 and larger than that of 2000-2001. Because of the large differences among surveys, it is diffcult to explore long-term change in macrobenthos abundance.However, that abundance has fluctuated within a certain range during various periods in the two seas.

Table 4 Comparison of species number and abundance of macrobenthos between Yellow and East China Seas during different periods
4.2 Relationship between macrobenthic communities and environmental factors in Yellow and East China Seas
In terms of the spatial distribution of the macrobenthic communities, except for summer, the number of macrobenthos in the Yellow Sea was significantly larger than that in the East China Sea.This is related to the environments of the seas,including sediment type and water mass.
First, the sediment types in these two seas are different. Seaf l oor sediment in the Yellow Sea is siltbased, whose particles are coarser than those of the sand-based sediment in the East China Sea (Wang,2013). The impact of sediment type on themacrobenthos distribution has two main two aspects.One is the availability of foods. That is, the pore size of sediment with fi ne particles is small, and nutrients in the sediment are not easily used by macrobenthos,leading to less food for macrobenthos than that in the sediment with coarse particles. In addition, there is various habitat heterogeneity in different types of sediments. The spatial heterogeneity of sediments with coarse particles is greater than that of sediments with fi ne particles. Strong spatial heterogeneity can not only provide diverse benthic habitats suitable for various types of benthic organisms in which to live,but also provide shelter from predators for such organisms. Therefore, species number, abundance and diversity of benthic organisms increase with the proportion of coarse gravels in sediments, according to Long and Lewis (1987). This distribution of benthic organisms has been confi rmed in other studies.Benthic biodiversity in sandy mud and muddy sand was greater than those in sand or clay (Zhao and Wang, 2000). The various sediment types in the Yellow and East China Seas would affect the ecological types, nutritional characteristics, and community structure of benthic organisms (Liu et al.,1986). Sediment type would inf l uence the abundance of benthic organisms, which is relatively low in sediment with fi ne particles (Jayaraj et al., 2008). At depths >40 m, macrobenthos abundances in medium and coarse sand sediments were 25% higher than in other sediment types (Coleman et al., 2007).
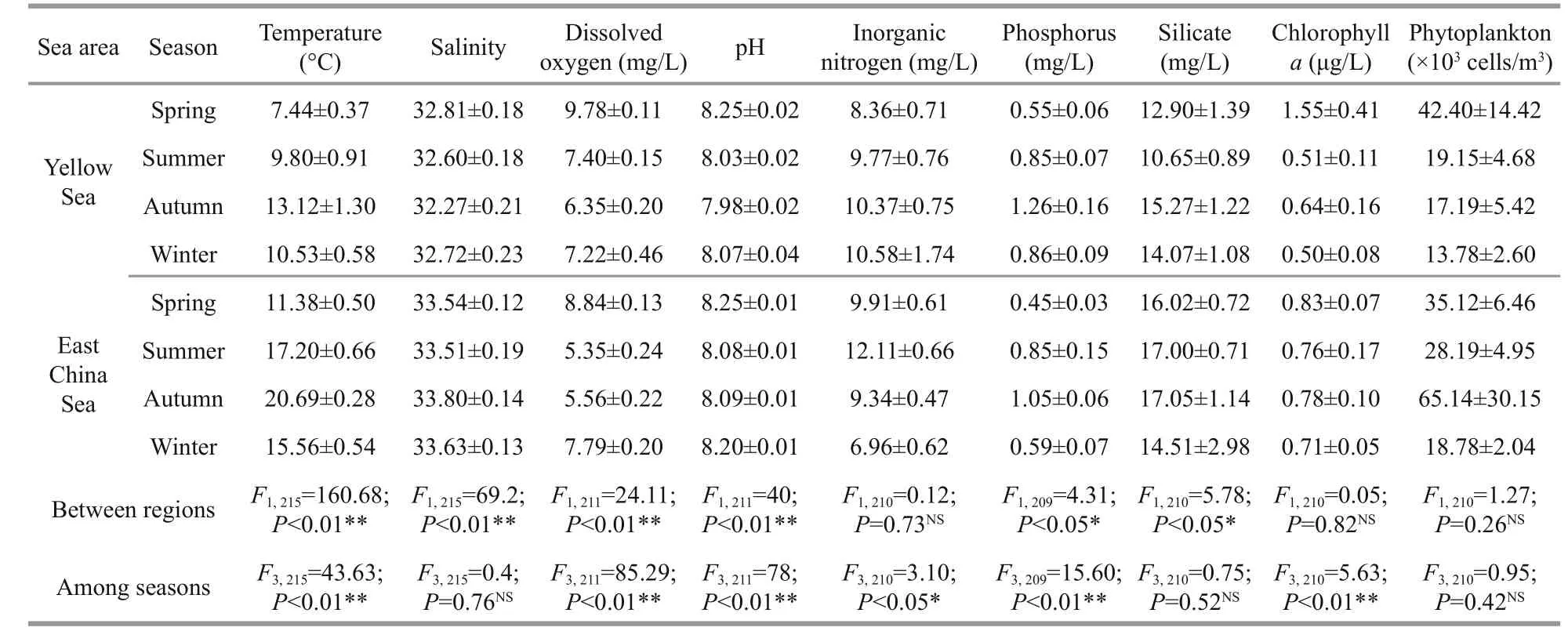
Table 5 Seasonal and regional variation of environmental factors in Yellow and East China Seas
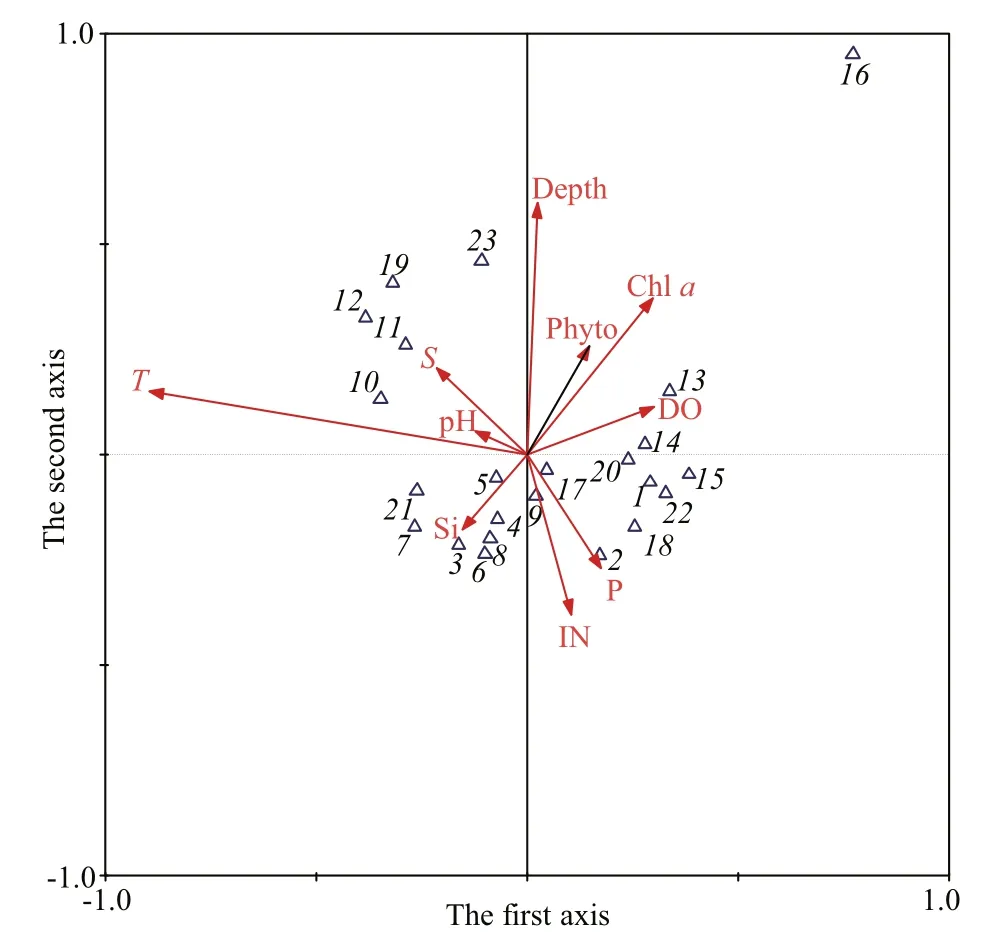
Fig.4 CCA ordination diagrams of macrobenthos and environmental factors in Yellow and East China SeasT: temperature;S: salinity; pH: degree of acidity or alkalinity;DO: dissolved oxygen; IN: inorganic nitrogen; P: phosphorus; Si:silicate; Chla: Chlorophylla; Phyto: phytoplankton; 1:Melinna cristata; 2:Terebellides stroemii; 3:Aglaophamus dibranchis; 4:Lumbrineris heteropoda; 5:Lumbrineris cruzensis; 6:Lumbrinerissp.; 7:Dasybranchus caducus; 8:Notomastus latericeus; 9:Ophelina acuminate; 10:Dentalium octangulatum; 11:Cavolinia uncinata; 12:Cavolinia longirostris; 13:Nucula(Leionucula)nipponica; 14:Nucula(Leionucula)tenuis; 15:Thyasira tokunagai;16:Raetellops pulchella; 17:Ampeliscasp.; 18:Eocuma lata;19:Callianassasp. 20:Ophiura sarsii vadicola; 21:Apionsoma trichocephala; 22:Portlandia japonica; 23:Cavolinia tridentate.
Second, the relatively complex hydrologic environments of the two seas (Table 5) could determine the ecological groups of macrobenthos.The deep water of the Yellow Sea (depth >40-50 m)is in its central part, where bottom water temperature is low and stable year-round. Dominant macrobenthos in this area areThyasiratokunagai,Portlandiajaponica,Ophiura sarsii vadicola, andMelinna cristata. The species composition of macrobenthic organisms there was almost constant between the 1950s and 1990s (Zhang, 2012). Because hydrologic conditions in continental-shelf waters of the East China Sea (open sea area distant from the 50-60 m isobath) is affected by the Kuroshio Current, bottom water temperature and salinity on the west side of this area are high year-round, and the dominant macrobenthos prefer high temperature and salinity.These includeDentaliumoctangulatum,Cavoliniasp., andCallianassasp. In contrast, the shallow coastal waters of the East China and Yellow Seas(within the 40-50 m isobath) are mainly affected by the continental climate and river runoff, which induce clear seasonal changes of temperature and salinity.Therefore, dominant macrobenthos in this region are eurythermal and euryhaline species, such asHeromastusfi liforms,Notomastuslatericeus,CapitellacapitataandAmpeliscasp. The complex hydrologic conditions in the two seas are undoubtedly important in determining the composition of benthic biota and differences in properties (Liu et al., 1986).CCA showed that temperature and water depth were the main environmental effects on the distribution of macrobenthos, revealing that with the action of different water masses, changes in bottom water temperature of the two seas inf l uenced the composition and distribution of benthic organisms. Biomass and the secondary productivity of macrobenthos in the East China Sea were significantly affected by water temperature and depth (Li et al., 2005b, c). In addition,owing to the impact of the YSCM, biomass and secondary productivity of the macrobenthos were strongly affected by water temperature (Li et al.,2005c). Water depth, temperature and median particle size in the sediment were important inf l uences on the distribution of macrobenthos in the southern Yellow Sea (Zhang, 2012). Similar patterns were found in plankton. For example, the species composition and distributions of planktonic bacteria, phytoplankton and zooplankton were all greatly shaped by water masses in the two seas (Li et al., 2006; Chen et al.,2011; Gao et al., 2013).
5 SUMMARY
As the world’s most important marginal seas, the East China and Yellow Seas have enormous pressures from rapid economic development. However, because of the high cost of marine ecological surveys, largescale spatiotemporal variations of macrobenthos communities have been unclear. Therefore, we conducted surveys during four cruises to show seasonal and spatial variations of macrobenthic communities in the Yellow and East China seas. In this study, we found the following: i) a total of 562 macrobenthic species were identified, with polychaetes and mollusks accounting for 67% of total species, with the dominant ones bivalve mollusks and small-sized polychaetes; ii) based on the two-way ANOVA and two-factor community similarity analysis, there were significant spatiotemporal variations of macrobenthic community, such as seasonal variations in number of species, density and diversity index, and regional differences of biomass and density; iii) based on relationships between macrobenthos and their living environments, sediment properties and water mass characteristics in the two seas were speculated to be the main causes of regional differences.
6 ACKNOWLEDGEMENT
We acknowledge HU Yuemei, WANG Bin, and JIN Haiyan for their assistance in the laboratory and fi eld.
 Journal of Oceanology and Limnology2018年3期
Journal of Oceanology and Limnology2018年3期
- Journal of Oceanology and Limnology的其它文章
- Response of the North Pacific Oscillation to global warming in the models of the Intergovernmental Panel on Climate Change Fourth Assessment Report*
- Effect of mesoscale wind stress-SST coupling on the Kuroshio extension jet*
- Surface diurnal warming in the East China Sea derived from satellite remote sensing*
- Cross-shelf transport induced by coastal trapped waves along the coast of East China Sea*
- Observations of near-inertial waves induced by parametric subharmonic instability*
- Seasonal variation and modal content ofinternal tides in the northern South China Sea*