
Palaeoenvironmental and biostratigraphic implications of microbial mat-related structures:Examples from the modern Gulf of Cambay and the Precambrian Vindhyan Basin, India
2014-09-27 02:35:54SantanuBanerjeeSubirSarkarPatrickEriksson
Journal of Palaeogeography 2014年2期
Santanu Banerjee , Subir Sarkar, Patrick G.Eriksson
1.Department of Earth Sciences, Indian Institute of Technology Bombay, Powai, Mumbai-400076, India 2.Department of Geological Sciences, Jadavpur University, Kolkata-700032, India 3.Department of Geology, University of Pretoria, Pretoria 0002, South Africa *
1 lntroduction
Microbial mat structures are well known from carbonates while microbial mat influence on clastic sediments has received increasing attention in recent years.Microbes impart unusual cohesiveness to clastic sediments and form a wide variety of structures, commonly known as microbially-induced sedimentary structures (MISS: Noffkeet al.,2001a)or microbial mat-related structures (MRS: Erikssonet al., 2010).We choose to use the latter term here, for its broader genetic connotations, but we also note that essentially the terms are analogues.In general, the palaeogeographic implications of mat-related structures from clastic sedimentary settings are poorly explored notwithstanding a good number of publications in the last decade from Precambrian deposits (Hagadorn and Bottjer, 1997; Noffkeet al., 2003; Banerjee and Jeevankumar, 2005; Parizotet al.,2005; Sarkaret al., 2005, 2006, 2008; Boseet al., 2007;Bouougri and Porada, 2007; Sarkar and Banerjee, 2007;Banerjeeet al., 2010; Erikssonet al., 2010, 2012; Noffke,2010; Tanget al., 2011; Lan and Chen, 2012, 2013; and many others).While uninterrupted mat growth was characteristic of Precambrian seas, extensive microbial mats may develop on modern hypersaline coasts.Study of modern MRS provides useful insights for understanding the origin of the wide spectrum of non-actualistic sedimentary features in Precambrian sedimentary rocks which are related to unusual cohesiveness of the original sediments (Gerdeset al., 1993, 1994; Noffkeet al., 2003; Gerdes, 2007; Bose and Chafetz, 2009, 2011; Banerjeeet al., 2010).Although a few studies have reported distribution patterns of MRS in modern coastal settings (Noffke, 2000; Noffkeet al.,2001b; Bose and Chafetz, 2009, 2011; Bouougri and Porada, 2012), the wide variability of MRS in peritidal settings needs further investigation.Thorough understanding regarding the distribution of MRS in modern environments is crucial for high-resolution palaeogeographic interpretations of Precambrian shallow marine siliciclastic successions.Furthermore, MRS may often be confused for burrows, tracks and trails found on bed surfaces, particularly those which may occur in Pre-Ediacaran rocks.Microbial mats may often assume a discoidal geometry in appearance on sediment surfaces and the relics of cm-scale ‘discoidal microbial colonies’ (DMC)may also resemble medusoidtype Ediacaran fossils (Gerdeset al., 1994; Grazhdankin and Gerdes, 2007; Banerjeeet al., 2010; Banerjee, 2012),but which create problems in that they may occur in considerably older rocks.
This paper presents the distribution pattern of MRS in modern coastal plains of the Gulf of Cambay on the western coast of India and discusses its palaeogeographic implications.Distribution of MRS in the modern environment is compared with those occurring in the ~1.6 Ga,shallow marine Chorhat Sandstone of the Vindhyan Basin in central India to highlight the usefulness of MRS for high-resolution palaeogeographic interpretations.MRS resembling metazoan traces and Ediacaran medusoids are presented from the >1 Ga Vindhyan Supergroup and these features are compared with the modern Gulf of Cambay analogues.
2 Geological background
2.1 Modern coastal environment of the Gulf of Cambay
Extensive microbial mat cover develops on siliciclastic sediments deposited on hypersaline intertidal and supratidal flats of the Gulf of Cambay on the western coast of India (Figure 1A).The N-S oriented Gulf of Cambay is macrotidal in nature, and the semidiurnal tidal range often exceeds 10 m in places (Figure 1A; Sahaet al., 2010).The western flank of the Gulf of Cambay is typified by carbonate sedimentation while the eastern flank of the gulf is characterized by siliciclastic deposition in several tide-dominated estuaries and tidal flats.The study area,the eastern flank of the Gulf of Cambay, witnesses heavy sediment discharge during the time of monsoonal rainfall between June and September and it remains dry for the rest of the year.The E-W oriented Narmada estuary forms tidal sand bar, sand flat, mixed flat, mud flat and salt marsh facies zones (Sahaet al., 2010).The study area represents an estuarine tidal flat as per the classification of Fan (2013)near the mouth of the Narmada estuary (Figure 1A).
2.2 Precambrian Vindhyan Supergroup, central India
The Vindhyan Supergroup in central India preservesan upto 4.5 km thick, undeformed and unmetamorphosed sedimentary succession (Figure 1B).A basin-wide unconformity divides the Vindhyan Supergroup into two units.The older unit, the Lower Vindhyan (also known as the Semri Group), consists of carbonate, sandstone, shale and volcaniclastics (Boseet al., 1997); while the younger unit,the Upper Vindhyan, consists predominantly of siliciclastics with minor carbonate rocks (Boseet al., 2001).The oldest rock dated from the supergroup is slightly older than 1.6 Ga (Rasmussenet al., 2002; Gopalanet al., 2013).The upper age limit of the supergroup remains controversial because of the presence of possible Ediacaran fossils(see Venkatachalaet al., 1996)and an ~0.6 Ga Sr isotope age is determined for the Bhander Limestone near the top of the sedimentary succession (Rayet al., 2003).Recent radiometric and palaeomagnetic evidence, however,clearly establishes that the entire Vindhyan Supergroup is older than 1 Ga (Maloneet al., 2008; Basu and Brickford,2013; Gopalanet al., 2013; Venkateshwarlu and Rao,2013).The Vindhyan Supergroup is dominated by shallow marine sedimentation in inner shelf, shoreface and coastal environments barring a few aeolian and outer shelf deposits.Alternations between transgressive systems tract(TST)and highstand systems tract (HST)cycles comprise the basic sequence stratigraphic architecture (Boseet al.,2001).The sandstones make up the HSTs while shale members of outer to inner shelf origin comprise the TSTs.
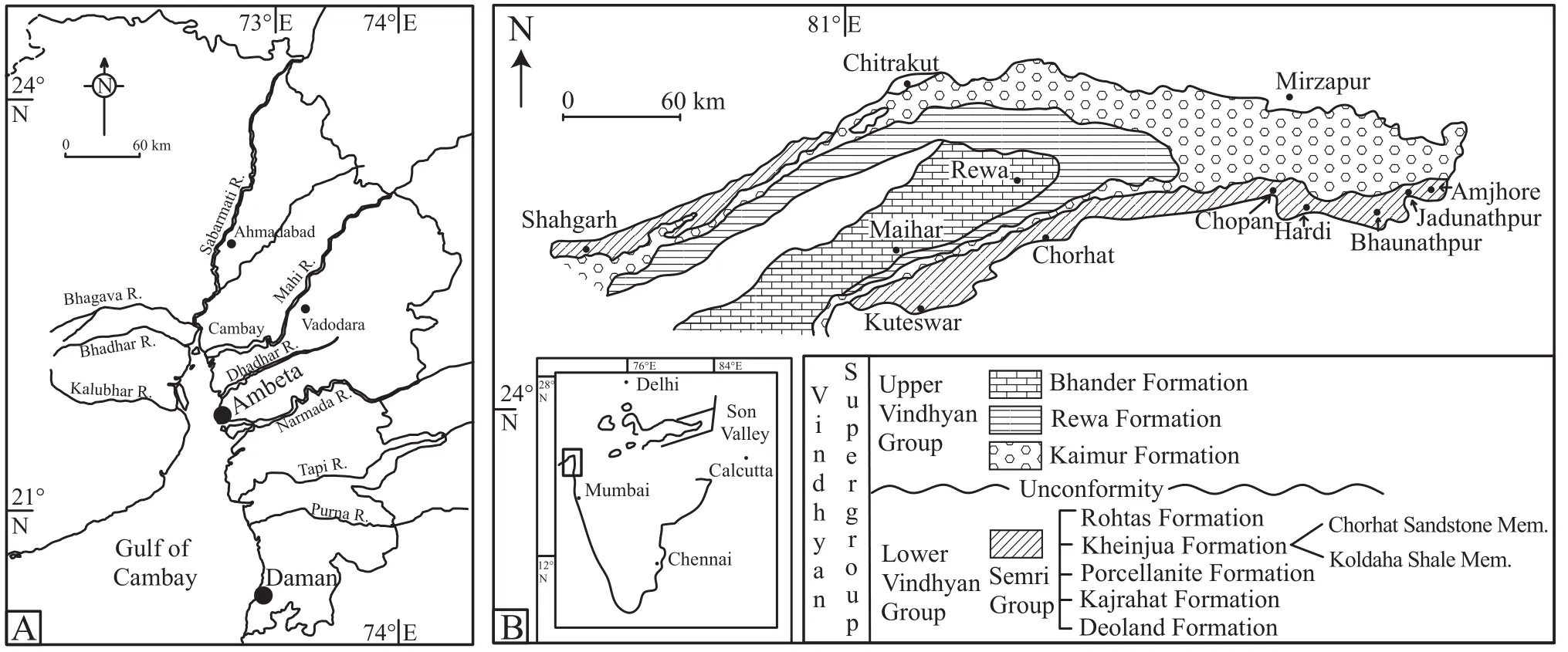
Figure 1 A-Map showing coastal areas around the Gulf of Cambay.The study area is close to Ambeta; B-Geological map showing outcrops of the Precambrian Vindhyan Supergroup along Son Valley with necessary stratigraphic elaborations.Map of India is within inset (Gulf of Cambay is marked by a rectangle).Mem.= Member.
This paper reports MRS features primarily from the~1.6 Ga Chorhat Sandstone of the Kheinjua Formation in the Lower Vindhyan, which is bounded by the Koldaha Shale at the base and the Rampur Shale above (Figure 2; Banerjee, 1997).The facies succession of the Chorhat Sandstone represents a shallow shelf bordered by a coastal aeolian flat.Sarkaret al.(2006)reported a subtidal to intertidal transition in the Chorhat Sandstone.The subtidal portion of the Chorhat Sandstone is constituted by sandy tempestites (average 35-40 cm thick), internally characterized by hummocky cross-stratifications intercalated with siltstones (Figure 2).The sandstone beds are frequently amalgamated upwards and are wavy laminated to quasi-planar laminated.The intertidal portion (average thickness 12 m)comprises well-sorted, fine-grained,wave-rippled sandstone with emergence features.The sandstones are well sorted and are completely devoid of mud particles.Very well sorted aeolian sheet sandstones,representing supratidal wind-blown sand sheets often occur on top of the intertidal facies.The MRS features occur within the subtidal and the intertidal facies (see Sarkaret al., 2006 for details).This paper reports MRS from the intertidal zone and compares them with modern examples from the Gulf of Cambay.
3 Distribution of MRS in the chosen modern environment
MRS are documented from four workable zones in the study area,viz., the lower intertidal, upper intertidal, lower supratidal and upper supratidal zones (Figure 3).The tidalzones are defined using the classification of Noffke (2010).The area located between the neap water line and the average low water line is the ‘lower intertidal zone’ and the area located between the average low water line and average high water line represents the ‘upper intertidal zone’.The‘lower supratidal zone’ demarcates the area between the average high water line and spring high water line, while the ‘upper supratidal zone’ occurs above the spring water line.The present study was conducted during the pre-monsoon period covering a complete spring-neap hemi-cycle.The predominantly muddy coastline remains inaccessible during the post-monsoon period.Hypersalinity across vast areas of the intertidal and supratidal flats of the study area allows profuse microbial mat growth (Figure 4A).
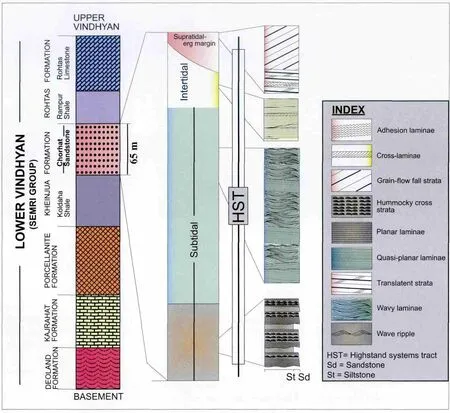
Figure 2 Stratigraphic framework of the Chorhat Sandstone, also showing overall facies succession.
3.1 MRS in the lower intertidal zone
MRS in the entire intertidal zone reflect the growth of microbial mats on sandy surfaces, decomposition of the mat cover, as well as physical destruction of the mat layer by waves and currents.The microbial mat cover is comparatively thicker in the shallow pools on the intertidal zone and within the troughs of ripples (Figures 3, 4B, 4C).MRS are relatively less diversified in the lower intertidal zone compared to the other zones as the mat cover is frequently destroyed there by relatively strong currents.However, wrinkle structures, patchy ripples and sieve-like surfaces may form in places (Figures 4D, 5A, 5B).Positive-relief wrinkle structures remain confined within shallow pools of local-ized mat growth (Figure 4D).On rippled surfaces the wrinkle structure mostly occupies the troughs, while the crests tend to be devoid of this feature.Small, linear positive relief wrinkles often form at a high angle to the ripple crests (Figure 5A).The wrinkles are distinguished from ladder-back ripples by their irregular, wavy nature and very low relief.These wrinkles are formed by the action of gentle, late stage currents on microbial mat-covered sands.Sieve-like surfaces, which represent numerous shallow holes bearing the impressions of gas bubbles, occur locally (Figure 5B).These are formed by the escape of gases through the mat cover.Sieve-like surfaces occur in local shallow pools of water or in the troughs of ripples and are analogous toKinneyiastructures (Figures 4C, 5B).The cohesive microbial mat forms a protective cover in the intertidal zone.However, relatively strong currents may tear away a portion of the mat cover and secondary ripples may form in these patches,which are known as patchy ripples (Sarkaret al., 2006).
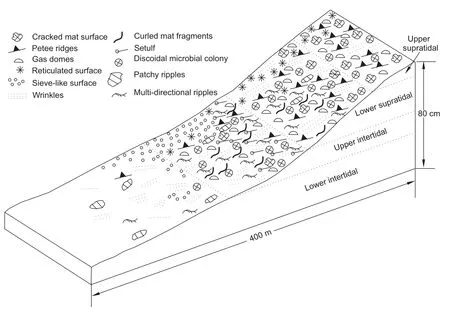
Figure 3 Schematic distribution of microbial mat-related structures (MRS)within the intertidal and supratidal zones of the Gulf of Cambay study area.
3.2 MRS in the upper intertidal zone
MRS features are more diversified in the upper intertidal zone as reduction in current strength mitigates physical destruction of the mat cover.The mat layer is often modified as it undergoes shrinkage during exposure.In addition to the features present in the lower intertidal zone, rolled-up mat fragments, gas domes, setulfs, multidirectional ripple marks and disc-shaped microbial colonies are formed in this zone (Figures 3, 5C-5F).Petee ridges and cracked mat surfaces are also found locally.Patches of secondary ripples often occur in a different direction completely surrounded by a primary ripple train.The boundary between the original rippled surface and reworked patches is sharply defined (Figure 5C).The feature is also known as multi-directional ripples (Noffke,1998; Noffkeet al., 2001a).Gas domes often form on mat surfaces by the escape of gases from decomposed mats underneath (Figure 5D).Joining of several gas domes may lead to elongated to irregular outlines.Gas domes preferentially occupy the troughs, while the crests of ripples may be devoid of these features (Figure 5D).The gas domes are distinguished from petee ridges by the hollow internal structures in the former (Bose and Chafetz,2011).Sands deposited within the intertidal zone attain enough cohesion from the mat cover, to form curled and rolled-up mat fragments, because of shrinkage.Some of the rolled-up mat fragments may be cigar-shaped (Figure 5E).The feature preferentially occupies local topographic highs which are prone to frequent exposure.Inverted fl ute cast-like features, known as setulfs, occur profusely close to the boundary between upper intertidal and lower supratidal zones.Although Friedman and Sanders (1974)described the feature from a modern tidal fl at, the origin of the features remained uncertain for a long time until Sarkaret al.(2011)provided examples of setulfs from both modern environments and ancient deposits and discussed its palaeogeographic implications.Forming against some minute obstacles like fecal pellets, tiny shell fragments or coarser sediment particles, setulfs may appear as tiny elongated ridges, the gentler sides of which indicate the downwind direction (Figure 5F).However,setulfs are very uncommon in the rock record because of a very poor preservation potential.

Figure 4 A-A fi eld photograph showing a portion of the upper Gulf of Cambay intertidal zone in the foreground.Hypersalinity in the intertidal zone (see white salt deposits)allows profuse growth of microbial mats; B-Thick mat growth in shallow pools of the intertidal zone showing gas bubbles (photo width = 35 cm); C-Microbial mat growth in ripple troughs (indicated by an arrow, photo width = 65 cm); D-Wrinkle structure (marked by an arrow)formed in a shallow pool of water (photo width = 45 cm).
3.3 MRS in the lower supratidal zone
Wrinkle structures, patchy ripples, multi-directional ripples, setulfs, gas domes, petee ridges, reticulated surfaces, disc-shaped microbial colonies and cracked mat surfaces are well developed within the lower supratidal zone(Figure 6A-6H).An increased number of petee ridges, gas domes and cracked mat surfaces distinguishes this zone from the upper intertidal zone (Figure 3).Reduced physical destruction of the mat by considerably weaker currents and modifi cation of the mat by desiccation cause a wide variability in MRS in the lower supratidal zone.Wrinkle structures of various morphologies are common (Figure 6A, 6B).Shrinkage of the mat cover, preferably in local zones of thicker mat growth, may form irregular wrinkles in patches, in association with cracked mat surfaces(Figure 6A).Gas domes (diameter ranging from 1.5 cm to 2.9 cm)occur frequently on wrinkled surfaces; the dome surfaces often exhibit exactly similar wrinkles as to those present in the background mat (Figure 6B).In places, the gas domes may exhibit radial cracks (Figure 6C).The cracks originate from the centre of the dome and propagate close to its flanks.Occasionally, several domes are partially merged to exhibit a lumpy appearance on the mat surface (Figure 6D).Gas domes are formed by the escape of gases from underneath the mat cover (Schieber, 2004;
see also Bose and Chafetz, 2011).Depending on the cohesiveness and thickness of the mat the domes may exhibit either radial cracks and/ or wrinkle features.Those formed in relatively thick and tough leathery mat surfaces retain the original wrinkles while doming.A thin but cohesive mat generates cracks during escape of gases from beneath.

Figure 5 Field photographs showing MRS in the Gulf of Cambay intertidal zone.A-Linear positive-relief wrinkles in ripple troughs(cover of pen = 5 cm); B- Sieve-like surfaces on shallow pools of the intertidal zone marked by an arrow (coin diameter = 2.3 cm);C-Multi-directional ripples (length of Swiss knife = 9 cm); D-Gas domes in ripple troughs marked by arrow (match-stick length =4.5 cm), merging of several gas domes results in elongated appearance in places; E-Rolled-up and curled mat fragments (marker pen length = 13.5 cm); F-Swarms of setulfs on a bed surface, wind direction from right to left (pen length = 14.5 cm).

Figure 6 Field photographs showing MRS in the Gulf of Cambay supratidal zone.A-Patches of wrinkles (marked by red arrow)associated with cracked mat surface (blue arrow); B-Wrinkles on gas domes (arrow)and in the background; C-A gas dome (arrow)bearing radial cracks; D-Gas domes showing lumpy appearance; E, F-Petee ridges; G-Reticulated surface; H-Cracked mat surface(coin diameter = 2.3 cm, length of Swiss knife = 9 cm).
3.4 MRS in the upper supratidal zone
The upper supratidal zone is characterized by an abundance of petee ridges, gas domes, cracked mat surfaces,wrinkled structures and reticulated surfaces (Figure 3).Setulfs, patchy ripples and multi-directional ripples are completely absent in this zone.Petee ridges in the supratidal flat exhibit different morphologies and these may appear as spherical, elliptical and elongated forms on the mat surface (Figure 6E, 6F).The widths of the petee ridges vary from a few millimeters to several centimeters.Petee ridges of different varieties reflect upward movement of gases and sediments through cracks in the mat cover.Mat surfaces may exhibit a reticulated petee-like pattern formed by cross-cutting, thread-like, linear bulges which form miniature domes and raised polygons (Figure 6G).Dried and desiccated mats frequently exhibit cracks (Figure 6H).
4 Discoidal microbial colonies (DMC)in a modern environment
A special variety of MRS, termed here ‘discoidal microbial colony’ (DMC)encompasses disc-shaped, cm-scale benthic microbial colonies that exhibit internally radial,concentric, reticulate or wrinkle features (Figure 7A-7H).The feature is least common in the lower intertidal zone and is most abundant in the upper supratidal zone (Figure 3).The most common variety of DMC in the intertidal zone includes partially collapsed, featureless discs (Figure 7A).The feature has a very sharp outline with a relief varying from a few millimeters to less than one centimeters.Figure 7B represents a DMC with central flat-lying portion of ~4 cm diameter and a gently sloping outer zone with radial ridges.DMC may exhibit a prominent groove which divides it into two prominent lobes (Figure 7C).Close observation reveals several miniature domes on both sides of the groove.The last two varieties are scattered throughout the mat-covered supratidal zone.
In the supratidal zone DMC preferably occurs in the central sunken portion of desiccation-cracked polygons.Occasionally, DMC may exhibit a relatively high relief(~1 cm)with several lobes in the inner portion and radial features near the outer portion (Figure 7D).A variety of the feature is represented by a low-relief DMC with reticulate inner part, radial ridges in the outer part and relatively irregular margin (Figure 7E).DMC with prominent internal wrinkles is very common in the entire supratidal zone (Figure 7F).The relief of the discs is low (<5 mm)and diameters of the discs vary between 6.5 cm and 7.4 cm.Many poorly to moderately formed discs often pass laterally to wrinkle structures (Figure 7F).A low-relief DMC in the intertidal zone exhibits a reticulate internal pattern and is devoid of a sharp outline (Figure 7G).The last variety of DMC exhibits concentric bands of alternating light and dark zones (Figure 7H).Diameters of this DMC vary between 6.6 cm and 7.5 cm and widths of the bands vary between 3 and 4 mm.This variety of DMC is also known as fairy-ring structure (see Gerdeset al., 1994;Gerdes, 2007).
5 MRS from the Precambrian Vindhyan Supergroup
Excellent preservation of MRS has already been reported from both subtidal and intertidal segments of the Chorhat Sandstone (Banerjee, 1997; Sarkaret al., 2004,2005, 2006; Sarkar and Banerjee, 2007; Boseet al., 2007).The intertidal segment of the Chorhat Sandstone exhibits a wide spectrum of MRS related to growth, physical destruction (by waves and currents)and decomposition of microbial mats.While the bottom-most part of the intertidal segment is marked by the occurrence of minor wrinkle structures and sieve-like surfaces, a gradual increase in diversity and abundance is recorded toward the upper part of the intertidal segment.
The lower part of the intertidal segment of the Chorhat Sandstone is dominated by wrinkle structures, sieve-like surfaces, gas domes, patchy ripples, multi-directional ripples, mat chips and rolled-up mat fragments (Figure 8A-8F).Wrinkles tend to occur only in the troughs of ripples while the crests are mostly completely devoid of these(Figure 8A).Closely-spaced, tiny holes on sandstone bed surfaces (also known asKinneyia, see Walcott, 1914)are analogous to the sieve-like surfaces described from the modern environment (compare Figure 8B with Figure 5B).Gas domes, occasionally collapsed at their central parts, are abundant on a few Chorhat sandstone bed surfaces (Figure 8C).Discrete sets of ripples may often occur in patches on bed surfaces (Figure 8D, 8E, 8F).The preservation of a single train of ripples, in several separate places may also be described as patchy (Figure 8D).The
margins of the patches have unusually sharp and jagged outlines and the bed surfaces between the patches may be planed off (Figure 7D).It is inferred that microbial mat formed a protective cover on rippled surfaces.The mat cover was torn away from most places, and ripples could survive only under the remnant mat layer, giving rise to these patchy ripples.A first generation ripple train may be reworked by a second generation ripple, in patches, with a different average crest orientation (Figure 8E, 8F).The primary ripples could survive in places where they were covered by a microbial mat and secondary ripples could form in places where the mat cover was torn away by current action.The feature is known as multi-directional ripples.Similar features are recorded from the upper intertidal and lower supratidal zones in the modern Gulf of Cambay environment (compare Figure 5C with Figure 8E,8F).Petee ridges may occur very locally in the lower intertidal segment of the Chorhat Sandstone, but these remain confined within ripple troughs (Figure 8G).Rolled-up mat fragments (Figure 8H)present in the Chorhat Sandstone are comparable to those occurring in the modern environment (see Figure 5E).

Figure 7 Discoidal microbial colonies (DMC)of various types (coin diameter = 2.3 cm, lens cap diameter = 6 cm; see text for detailed description).

Figure 8 MRS in the lower part of the intertidal segment of the Chorhat Sandstone.A-Wrinkle structures within troughs of ripples(arrow); B-Kinneyia on bed surfaces; C-Gas domes (marked by arrow)with circular outline on a sandstone bed surface; D-Patchy ripples; E, F-Multi-directional ripples; G-Petee ridges in troughs of ripples; H-Rolled-up mat fragments (arrow)(length of Swiss knife = 9 cm, coin diameter in B = 2.3 cm, in C = 2.1 cm, pen length in D = 14 cm, tape in F = 40 cm).
The upper part of the intertidal segment of the Chorhat Sandstone is dominated by petee ridges, gas domes and inferred cracked mat surfaces (preserved as sand cracks)besides the occurrence of wrinkle structures,Kinneyiaand occasional multi-directional ripples and patchy ripples(Figure 9A-9H).Petee ridges may be of diverse shapes and sizes.The widths of the ridges may vary from less than 1 cm to 1.2 cm (Figure 9A, 9B).Mat-covered sands in the Chorhat palaeoenvironment became cohesive enough to form cracks of different morphologies which were filled by overlying sediments (Figure 9C-9F).More commonly the cracks exhibit cross-cutting patterns with triple junctions (Figure 9D), or a polygonal pattern (Figure 9E).Less commonly sinuous varieties of cracks may occupy the troughs of ripples (Figure 9F).These cracks are commonly known asManchuriophycus, which were probably formed by the localised desiccation of a thickened mat cover in ripple troughs (Pflueger, 1999).Manchuriophycus-type cracks are well known from the ~2.1 Ga Magaliesberg Formation (Parizotet al., 2005)and the ~2.9 Ga Mozaan Group of South Africa (Noffkeet al., 2003).The filled sinuous sand-cracks are analogous to petee ridges.However,in the former case the infilled sands are compositionally different, the cracks are mostly V-shaped and laminae are undisturbed in the vertical section.Petee ridges generally comprise sands from the bed on which they lie and they are associated with deformation of laminae in the vertical section, being also separated by concave upward segments between the ridges.Gas domes may be abundant on most Chorhat upper intertidal sandstone bed surfaces (Figure 8G).Merging of several gas domes may turn them into more elongated forms in places (Figure 8H).
In summary, MRS in the upper part of the intertidal segment of the Chorhat Sandstone are distinguished from those in the lower part by the abundance of gas domes,petee ridges and filled sand-cracks in the former.Multidirectional ripples, patchy ripples and rolled-up mat fragments are less common in the upper intertidal segment, but are abundant in the lower intertidal segment of the Chorhat Sandstone.Wrinkle structures and sieve-like surfaces occur in similar abundance in the entire intertidal segment.Both these features occur preferentially in the troughs and are absent on the crests of ripples and therefore suggest a very shallow water condition.
Banerjeeet al.(2010)reported many varieties of relics of DMC from the Vindhyan Supergroup and discussed their implications.A problematic feature on a wave rippled sandstone bed of the Chorhat Sandstone, with concentric arrangement of beads, may be best explained as a DMC (Figure 10A).The feature may relatively easily be confused with a feeding burrow of an organism, however the intact nature of the ripples rules out a burrow origin.Roughly circular, wrinkled masses with sharp outlines are occasionally found underneath the shelf-originated storm sandstone beds of the Bhander Formation (Figure 10B-10D).De (2003, 2006)considered these features as Ediacaran fossils.However, recent radiometric dating rules out any possible Ediacaran affinity for these features.The features should perhaps best be considered as disc-shaped microbial colonies, which reflect the effects of compaction in places.
6 Discussion
6.1 Palaeogeographic implications of the mat features
There have been very few investigations exploring the environmental affinities of specific groups of MRS in modern environments (Noffkeet al., 2001b; Bose and Chafetz, 2009, 2011; Banerjeeet al., 2010; Bouougri and Porada, 2012).MRS in ancient clastic sedimentary successions are most commonly documented from shallow marine littoral-supratidal environments (Schieberet al.,2007; Erikssonet al., 2010).Although the origin of microbial mats and to a degree also of MRS requires a gener-ally low-energy condition, combined with non-deposition(Gerdes, 2007), high-energy events are required for the formation of a variety of MRS including patchy ripples,multi-directional ripples, rolled-up mat fragments and mat chips.Also, MRS formed at one place may be transported to other palaeoenvironments.Erikssonet al.(2010)suggested three broad categories of MRS including, (1)those occurring in shallow marine littoral-supratidal settings,(2)those occurring in shallow subtidal to deep marine settings and (3)those not showing preferential relationship to specific palaeoenvironments.The distribution of MRS in a modern intertidal-supratidal environment is useful to establish the palaeoenvironmental affinity of these features in ancient shallow marine deposits.

Figure 10 A-A possible discoidal microbial colony (marked by arrow)with concentric internal features (coin diameter = 1.9 cm); B,C, D-Discoidal microbial colonies marked by arrows, in the Bhander Formation (length of bar in B = 4 cm, match-stick length in C =4.5 cm and coin diameter in D = 1.9 cm).
In the present study, comparison with the distribution of modern MRS identifies the lower part of the intertidal segment of the Chorhat Sandstone as the upper intertidal zone,while the upper part of the Chorhat intertidal segment represents deposition close to the high-tide line.The abundance of filled sand cracks, petee ridges and gas domes in the upper part of the Chorhat intertidal segment suggests prolonged exposure of the mat-covered sands.The bottom-most part of the intertidal segment of the Chorhat Sandstone represents the transition from the subtidal to the lower intertidal zone.Wrinkle structures are present throughout the modern intertidal and supratidal zones and are, therefore poor palaeoenvironmental indicators.Although rolled-up mat fragments are found in the upper intertidal to lower supratidal zone in modern environments,including the Gulf of Cambay, similar features may occur in the subtidal settings of ancient deposits; the action of storm currents on mat surfaces may generate rolled-up and curled mat fragments (Simonson and Carney, 1999).The wrinkle structures may occur all over the entire intertidal and supratidal zones.This feature is also reported from outer shelf (Banerjee and Jeevankumar, 2005)and deep marine environments (Schieberet al., 2007).Therefore,a group of MRS may show no preferential relationship to specific environments.It has been realized that wrinkle structures, rolled-up structures and patchy ripples can occur within a broad range of palaeogeographic settings,from supratidal to subtidal conditions (Schieberet al.,2007; Erikssonet al., 2010).However, petee ridges, sandcracks, gas domes, multi-directed ripple marks, reticulate surfaces, sieve-like surfaces and setulfs are confined to the shallowest parts of the marine basins, representing upper intertidal to supratidal conditions.
6.2 Biostratigraphic implications of MRS
Microbial mats and discoidal microbial colonies may form a wide variety of features which may resemble Ediacaran fossils, and these potentially confusing features may be found in considerably older rocks as well (Banerjeeet al., 2010).In the absence of precise radiometric dating such features may be mistaken for the Neoproterozoic fossils.Microbially originated sinuous cracks in rippled sandstones,Manchuriophycus(Figure 9F), are often confused with metazoan burrows (e.g., Kulkarni and Borkar,1996 and many others).Wrinkle structures are mistakenly considered to beProtopaleodictyonandSquamodictyon(Durandet al., 1994).The wrinkle structures presented from the modern coastal Cambay environment and the Precambrian Chorhat Sandstone appear similar to EdiacaranArumberia.Kumar and Pandey (2008)argued for an Ediacaran affinity of anArumberia-like feature in the Upper Vindhyan.Petee ridges (Figure 9A, 9B)are confused with horizontal burrows (Singh and Sinha, 2001).Gas domes with central depression (Figure 8C)or radial features may be confused with jellyfish impressions (Glaessner and Wade, 1966; Sun, 1986a, 1986b; Cruseet al., 1993; De,2003, 2006; Srivastava, 2012 and many others).
Disc-shaped fossils are the most common component in Ediacaran biota and are often loosely termed as ‘medusoids’ (Sun, 1986a, 1986b).However, there is no unanimity among researchers regarding the exact affinity of the disc-shaped fossils (MacGabhann, 2007 and references therein).These have been variously interpreted as discoidal benthic organisms including cnidarians, porifera and fungi (Glaessner, 1984; Fedonkin, 1990; Gehling, 1991;Narbonne, 2005).A microbial affinity of the Ediacaran medusoids is often discarded because of a lack of modern analogues.However, Grazhdankin and Gerdes (2007)reinterpreted some of the well-known Ediacaran fossils with concentric zonations (e.g.Paliella,CyclomedusaandEdiacaria)as DMC.They have demonstrated that the disc-shaped fossils are essentially made up of microscopic filaments and the concentric zonation in the fossil specimens are equivalent to the concentric rings observed in a certain variety of DMC in modern coastal environments,known as a ‘fairy ring’ feature (Figure 6H).This paper, for the first time reports various internal structures found in DMC.Concentric zoning and radially oriented sectors, the most significant macroscopic features of the DMC, have been observed in microbial colonies forming under laboratory conditions (Shapiro and Dworkin, 1997; Shapiro,1998; Matsushitaet al., 2004)and in natural environments in rare cases (Gerdeset al., 1993, 1994; Banerjeeet al.,2010).The laboratory experiments and the Gulf of Cambay examples of DMC provide useful modern analogues of ancient discoidal Ediacaran fossils.The wide variation in the internal structure of DMC may be mistaken as morphological diversity of the medusoid group of Ediacaran fossils and colonial fossils in pre-Ediacaran rocks.
7 Conclusions
1)The abundance of MRS increases from the lower intertidal zone to the upper intertidal zone, while they are more diverse in the upper intertidal and lower supratidal zones in the modern environment.
2)The association of MRS may be used for high-resolution palaeoenvironmental interpretations in Precambrian shallow marine successions.The upper part of the intertidal segment of the Chorhat Sandstone is distinguished from the lower part by the presence of abundant cracked mat surfaces, petee ridges and gas domes in the former, while the abundance of wrinkle structures,Kinneyia, rolled-up mat fragments, patchy ripples and multi-directional ripples remains the same in both parts.Comparison of the distribution pattern of MRS in the studied modern environment suggests that the lower part of the intertidal segment of the Chorhat Sandstone equates to the upper intertidal zone of the modern environment, while the upper part of the intertidal segment of the Chorhat Sandstone represents prolonged exposure close to the high-tide line.
3)Discoidal microbial colonies may exhibit diverse internal structures and the feature may be mistaken for Ediacaran medusoids in considerably older rocks.Petee ridges, filled sand-cracks, gas domes and curled mat fragments may similarly be misidentified as traces of advanced organisms in Pre-Ediacaran rocks.Such problematic features should be ignored for correlation of end-Precambrian sedimentary successions.
Banerjee, S., 1997.Facets of the Mesoproterozoic Semri sedimentation in Son Valley, India [Ph.D.thesis].Jadavpur University,Kolkata, 1-137.
Banerjee, S., 2012.Discoidal microbial colonies.International Jour-nal of Earth Science, 101(5): 1343.
Banerjee, S., Jeevankumar, S., 2005.Microbially originated wrinkle structures on sandstones and their stratigraphic context: Palaeoproterozoic Koldaha Shale, central India.Sedimentary Geology,176(1-2): 211-224.
Banerjee, S., Sarkar, S., Eriksson, P.G., Samanta, P., 2010.Microbially related structures in siliciclastic sediment resembling Ediacaran fossils: examples from India, ancient and modern.In: Seckbach J., Oren, A., (eds).Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems.Berlin: Springer-Verlag,111-129.
Basu, A., Bickford, M.E., 2013.Contributions of zircon U-Pb geochronology to understanding the volcanic and sedimentary history of some Purāna basins, India.Journal of Asian Earth Sciences,doi: http://dx.doi.org/10.1016/j.jseaes.2013.06.018 (in press).
Bose, P.K., Banerjee, S., Sarkar, S., 1997.Slope-controlled seismic deformation and tectonic framework of deposition of Koldaha Shale, India.Tectonophysics, 269(1-2): 151-169.
Bose, P.K., Sarkar, S., Banerjee, S., Chakraborty, S., 2007.Matrelated features from the Vindhyan Supergroup in central India.In: Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S., Sarkar,S., Catuneanu, O., Altermann, W., (eds).An Atlas of Microbial Mat Features Preserved within the Clastic Rock Record.Amsterdam: Elsevier, 181-188.
Bose, P.K., Sarkar, S., Chakraborty, S., Banerjee, S., 2001.Overview of the Meso- to Neoproterozoic evolution of the Vindhyan Basin,central India.Sedimentary Geology, 141-142: 395-419.
Bose, S., Chafetz, H.S., 2009.Topographic control on distribution of modern microbially induced sedimentary structures (MISS):A case study from Texas coast.Sedimentary Geology, 213(3-4):136-149.
Bose, S., Chafetz, H.S., 2011.Morphology and distribution of MISS: A comparison between modern siliciclastic and carbonate settings.In: Noffke, N., Chafetz, H., (eds).Microbial Mats in Siliciclastic Depositional Systems Through Time.SEPM Special Publications, 101: 3-14.
Bouougri, E.H., Porada, H., 2007.Mat-related features from the Neoproterozoic Tizi n-Taghatine Group, Anti-Atlas belt, Morocco.In: Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S., Sarkar,S., Altermann, W., Catuneanu, O., (eds).Atlas of Microbial Mat Features Preserved within the Clastic Rock Record.Amsterdam:Elsevier, 198-207.
Bouougri, E.H., Porada, H., 2012.Wind-induced mat deformation structures in recent tidal flats and sabkhas of SE-Tunisia and their significance for environmental interpretation of fossil structures.Sedimentary Geology, 263-264: 56-66.
Cruse, T., Harris, L.B., Rasmussen, B., 1993.Geological note: The discovery of Ediacaran trace and body fossils in the Stirling Range Formation, Western Australia: Implications for sedimentation and deformation during the ‘Pan-African’ orogenic cycle.Australian Journal of Earth Science, 40(3): 293-296.
De, C., 2003.Possible organisms similar to Ediacaran forms from the Bhander Group, Vindhyan Supergroup, Late Neoproterozoic of India.Journal of Asian Earth Science, 21(4): 387-395.
De, C., 2006.Ediacara fossil assemblage in the Upper Vindhyans of central India and its significance.Journal of Asian Earth Science,27(5): 660-683.
Durand, F.R., Lech, R.R., Tortello, M.F., 1994.Nuevas evidencias paleontológicas en el basamento Precámbrico-Cámbrico del noroeste argentino.Acta Geológica Leopoldensia, 39(2): 691-701.
Eriksson, P.G., Bartman, R., Catuneanu, O., Mazumder, R., Lenhardt, N., 2012.A case study of microbial mat-related features in coastal epeiric sandstones from the Palaeoproterozoic Pretoria Group (Transvaal Supergroup, Kaapvaal craton, South Africa);The effect of preservation (reflecting sequence stratigraphic models)on the relationship between mat features and inferred palaeoenvironment.Sedimentary Geology, 263/264: 67-75.
Eriksson, P.G., Sarkar, S., Banerjee, S., Porada, H., Catuneanu, O.,Samanta, P., 2010.Paleoenvironmental context of microbial matrelated structures in siliciclastic rocks: Examples from the Proterozoic of India and South Africa.In: Seckbach, J., Oren, A.,(eds).Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems.Berlin: Springer-Verlag, 73-108.
Fan, D.D., 2013.Marine sedimentology classifications, sedimentary features and facies associations of tidal flats.Journal of Palaeogeography, 2(1): 66-80.
Fedonkin, M.A., 1990.Systematic description of Vendian metazoan.In: Sokolov, B.S., Iwanoski, A.B., (eds).The Vendian System:Paleontology 1.Berlin: Springer-Verlag, 71-120.
Friedman, G.M., Sanders, J.E., 1974.Positive-relief bedforms on modern tidal-flat that resemble moulds of flutes and grooves: implications for geopetal criteria and for origin and classification of bedforms.Journal of Sedimentary Petrology, 44(1): 181-189.
Gehling, J.G., 1991.The case for Ediacaran fossil roots to the metazoan tree.Memoir of the Geological Society of India, 20: 181-224.
Gerdes, G., 2007.Structures left by modern microbial mats in their host sediments.In: Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S., Sarkar, S., Altermann, W., Catuneanu, O., (eds).Atlas of Microbial Mat Features Preserved within the Siliciclastic Rock Record.Amsterdam: Elsevier, 5-38.
Gerdes, G., Claes, M., Dunajtschik-Piewak, K., Riege, H., Krumbein, W.E., Reineck, H.E., 1993.Contribution of microbial mats to sedimentary surface structures.Facies, 29(1): 61-74.
Gerdes, G., Krumbein, W.E., Reineck, H.E., 1994.Microbial mats as architects of sedimentary surface structures.In: Krumbein, W.E., Paterson, D.M., Stal, L.J., (eds).Biostabilization of Sediments.Oldenburg: Bibliotheks und Informationssytem der Carl von Ossietzky Universit?t Oldenburg (BIS)-Verlag, 165-182.
Glaessner M.F., 1984.The Dawn of Animal Life: A Biohistorical Study.Cambridge: Cambridge University Press, 1-296.
Glaessner, M.F., Wade, M., 1966.The Late Precambrian fossils from the Ediacara, South Australia.Palaeontology, 9(4): 599-628.
Gopalan, K., Kumar, A., Kumar, S., Vijayagopal, B., 2013.Depositional history of the Upper Vindhyan succession, central India:Time constraints from Pb-Pb isochron ages of its carbonate components.Precambrian Research, 233: 108-117.
Grazhdankin, D., Gerdes, G., 2007.Ediacaran microbial colonies.Lethaia, 40(3): 201-210.
Hagadorn, J.W., Bottjer, D.J., 1997.Wrinkle structures: Microbially mediated sedimentary structures common in subtidal siliciclastic settings at the Proterozoic-Phanerozoic transition.Geology,25(11): 1047-1050.
Kulkarni, K.G., Borkar, V.D., 1996.Occurrence ofCochlichnusHitchcock in the Vindhyan Supergroup (Proterozoic)of Madhya Pradesh.Journal of the Geological Society of India, 47(6): 725-729.
Kumar, S., Pandey, S.K., 2008.Arumberiaand associated fossils from the Neoproterozoic Maihar Sandstone, Vindhyan Supergroup, Central India.Journal of the Palaeontological Society of India, 53(1): 83-97.
Lan, Z.W., Chen, Z.Q., 2012.Exceptionally preserved microbially induced sedimentary structures from the Ediacaran post-glacial successions in the Kimberley region, northwestern Australia.Precambrian Research, 200-203: 1-25.
Lan, Z.W., Chen, Z.Q., 2013.Proliferation of MISS-forming microbial mats after the late Neoproterozoic glaciations: Evidence from the Kimberley region, NW Australia.Precambrian Research, 224: 529-550.
MacGabhann, B.A., 2007.Discoidal fossils of the Ediacaran biota: A review of current understanding.In: Vickers-Rich, P., Komarower, P., (eds).The Rise and Fall of the Ediacaran Biota.Geological Society of London, Special Publications, 286: 297-313.
Malone, S.J., Meert, J.G., Banerjee, D.M., Pandit, M.K., Tamrat,E., Kamenov, G.D., Pradhan, V.R., Sohl, L.E., 2008.Paleomagnetism and detrital zircon geochronology of the Upper Vindhyan Sequence, Son Valley and Rajasthan, India: A ca.1000 Ma closure age for the Purana Basins? Precambrian Research,164(3-4): 137-159.
Matsushita, M., Hiramatsu, F., Kobayashi, N., Ozawa, T., Yamazaki,Y., Matsuyama, T., 2004.Colony formation in bacteria: Experiments and modelling.Biofilms, 1(4): 305-317.
Narbonne, G.M., 2005.The Ediacara biota: Neoproterozoic origin of animals and their ecosystems.Annual Review of Earth and Planetary Sciences, 33: 421-442.
Noffke, N., 1998.Multidirected ripple marks rising from biological and sedimentological processes in modern lower supratidal deposits (Mellum Island, southern North Sea).Geology, 26(10):879-882.
Noffke, N., 2000.Extensive microbial mats and their influences on the erosional and depositional dynamics of a siliciclastic cold water environment (Lower Arenigian, Montagne Noire, France).Sedimentary Geology, 136(3-4): 207-215.
Noffke, N., 2010.Geobiology: Microbial Mats in Sandy Deposits from the Archean Era to Today.Berlin: Springer-Verlag, 1-194.
Noffke, N., Gerdes, G., Klenke, T., Krumbein, W.E., 2001a.Microbially induced sedimentary structures: A new category within the classification of primary sedimentary structures.Journal of Sedimentary Research, 71(5): 649-656.
Noffke, N., Gerdes, G., Klenke, T., Krumbein, W.E., 2001b.Microbially induced sedimentary structures indicating climatological,hydrological and depositional conditions within recent and Pleistocene coastal facies zones (Southern Tunisia).Facies, 44(1):23-30.
Noffke, N., Hazen, R., Nhleko, N., 2003.Earth's earliest microbial mats in a siliciclastic marine environment (2.9 Ga Mozaan Group, South Africa).Geology, 31(8): 673-676.
Parizot, M., Eriksson, P.G., Aifa, T., Sarkar, S., Banerjee, S.,Catuneanu, O., Altermann, W., Bumby, A.J., Bordy, E.M.,van Rooy, J.L., Boshoff, A.J., 2005.Suspected Microbial matrelated crack-like sedimentary structures in the Paleoproterozoic Magaliesberg Formation sandstones, South Africa.Precambrian Research, 138(3-4): 274-296.
Pflueger, F., 1999.Matground structures and redox facies.Palaios,14(1): 25-39.
Rasmussen, B., Bose, P.K., Sarkar, S., Banerjee, S., Fletcher, I.R., McNaughton, N.J., 2002.1.6 Ga U-Pb zircon ages for the Chorhat Sandstone, lower Vindhyan, India: Possible implications for early evolution of animals.Geology, 30(2): 103-106.
Ray, J.S., Veizer, J., Davis, W.J., 2003.C, O, Sr and Pb isotope systematics of carbonate sequences of the Vindhyan Supergroup,India: Age, diagenesis, correlations and implications for global events.Precambrian Research, 121(1-2): 103-140.
Saha, S., Banerjee, S., Burley, S.D., Ghosh, A., Saraswati, P.K.,2010.The influence of flood basaltic source terrains on the efficiency of tectonic setting discrimination diagrams: An example from the Gulf of Khambhat, western India.Sedimentary Geology, 228(1-2): 1-13.
Sarkar, S., Banerjee, S., 2007.Some unusual and/or problematic features.In: Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee,S., Sarkar, S., Catuneanu, O., Altermann, W., (eds).An Atlas of Microbial Mat Features Preserved within the Clastic Rock Record.Amsterdam: Elsevier, 145-147.
Sarkar, S., Banerjee, S., Eriksson, P.G., 2004.Microbial mat features in sandstones illustrated.In: Eriksson, P.G., Altermann,W., Nelson, D.R., Mueller, W.U., Catuneanu, O., (eds).The Precambrian Earth: Tempos and Events.Amsterdam: Elsevier,673-675.
Sarkar, S., Banerjee, S., Eriksson, P.G., Catuneanu, O., 2005.Microbial mat control on siliciclastic Precambrian sequence stratigraphic architecture: Examples from India.Sedimentary Geology, 176(1-2): 195-209.
Sarkar, S., Banerjee, S., Samanta, P., Jeevankumar, S., 2006.Microbial mat-induced sedimentary structures in siliciclastic sediments: examples from the 1.6 Ga Chorhat Sandstone,Vindhyan Supergroup, M.P., India.Journal of Earth System Science, 115(1): 49-60.
Sarkar, S., Bose, P.K., Samanta, P., Sengupta, P., Eriksson, P.G.,2008.Microbial mat mediated structures in the Ediacaran Sonia Sandstone, Rajasthan, India, and their implications for Proterozoic sedimentation.Precambrian Research, 162(1-2):248-263.
Sarkar, S., Samanta, P., Altermann, W., 2011.Setulfs, modern and ancient: Formative mechanism, preservation bias and palaeoenvironmental implications.Sedimentary Geology, 238(1-2): 71-78.
Schieber, J., 2004.Microbial mats in the siliciclastic rock record:A summary of the diagnostic features.In: Eriksson, P.G.,Altermann, W., Nelson, D.R., Mueller, W.U., Catuneanu, O.,(eds).The Precambrian Earth: Tempos and Events.Amsterdam:Elsevier, 663-672.
Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S., Sarkar,S., Catuneanu, O., Altermann, W., 2007.Atlas of Microbial Mat Features Preserved Within the Siliciclastic Rock Record.Amsterdam: Elsevier, 1-311.
Shapiro, J.A., 1998.Thinking about bacterial populations as multicellular organisms.Annual Review of Microbiology, 52:81-104.
Shapiro, J.A., Dworkin, M., 1997.Bacteria as Multicellular Organisms.Oxford: Oxford University Press, 1-466.
Simonson, B.M., Carney, K.E., 1999.Roll-up structures: Evidence of in situ microbial mats in Late Archean deep shelf environments.Palaios, 14(1): 13-24.
Singh, S.P., Sinha, P.K., 2001.Vindhyan Supergroup of Bihar — An overview.In: Singh S.P., (ed).Precambrian Crustal Evolution and Mineralisation in India.Patna: South Asian Association of Economic Geologists, 107-125.
Srivastava, P., 2012.Morphodiversity, complexity and macroevolution: Revealed by the megascopic life of the Palaeo-Neoproterozoic Vindhyan Supergroup, India.Geological Society of London,Special Publications, 365(1): 247-262.
Sun, W.G., 1986a.Precambrian medusoids: TheCyclomedusa plexusandCyclomedusa-like pseudofossils.Precambrian Research,31(4): 325-360.
Sun, W.G., 1986b.Late Precambrian scyphozoan medusaMawsonites randellensissp.nov.and its significance in the Ediacaran metazoan assemblage, South Australia.Alcheringa: An Australasian Journal of Palaeontology, 10(3): 169-181.
Tang, D.J., Shi, X.Y., Jiang, G., Wang, X.Q., 2011.Morphological association of microbially induced sedimentary structures(MISS)as a paleoenvironmental indicator: An example from the Proterozoic succession of the southern North China Platform.In:Noffke, N., Chafetz, H., (eds).Microbial Mats in Siliciclastic Depositional Systems Through Time.SEPM Special Publications, 101: 163-175.
Venkatachala, B.S., Sharma, M., Shukla, M., 1996.Age and life of the Vindhyans — facts and conjectures.Memoir of the Geological Society of India, 36: 137-165.
Venkateshwarlu, M., Rao, J.M., 2013.Palaeomagnetism of Bhander sediments from Bhopal Inlier, Vindhyan Supergroup.Journal of the Geological Society of India, 81(3): 330-336.
Walcott, C.D., 1914.Cambrian geology and palaeontology III No.2—Precambrian, Algonkian algal flora.Smithsonian Miscellaneous Collection, 64: 77-156.
 Journal of Palaeogeography2014年2期
Journal of Palaeogeography2014年2期
- Journal of Palaeogeography的其它文章
- Palinspastic reconstruction and geological evolution of Permian residual marine basins bordering China and Mongolia
- Dating recent sediments from the subaqueous Yangtze Delta and adjacent continental shelf, China
- Callovian-Oxfordian hecticoceratins from western India: Their biostratigraphic and palaeobiogeographic implications
- Restrictions to the application of ‘diagnostic’criteria for recognizing ancient seismites
- Jurassic sedimentary evolution of southern Junggar Basin: Implication for palaeoclimate changes in northern Xinjiang Uygur Autonomous Region, China
- Peritidal carbonate cycles induced by carbonate productivity variations: A conceptual model for an isolated Early Triassic greenhouse platform in South China